Индуцированные стволовые клетки

Индуцированные стволовые клетки (иСК) — стволовые клетки, полученные из каких-либо иных (соматических, репродуктивных или плюрипотентных) клеток путём эпигенетического перепрограммирования. В зависимости от степени дедифференцировки клетки при перепрограммировании различают: индуцированные тотипотентные, индуцированные плюрипотентные стволовые клетки (ИПСК) и получаемые так называемым прямым перепрограммированием или каким-либо иным способом индуцированные прогениторные (мультипотентные или унипотентные) стволовые клетки, иногда называемые также индуцированными соматическими стволовыми клетками (ИССК).
В настоящее время существует три пути перепрограммирования соматических клеток в плюрипотентные стволовые клетки:
- пересадка ядер, взятых из соматических клеток, в оплодотворенную яйцеклетку, из которой предварительно удалено ядро
- слияние соматических клеток с плюрипотентными стволовыми клетками;
- модификация соматической клетки, индуцирующая её превращение в стволовую клетку, с помощью: генетического материала, кодирующего белковые репрограммирующие факторы; рекомбинантных белков; микроРНК, синтетической самореплицирующейся полицистронной РНК и низкомолекулярных биологически активных веществ.
Содержание
- 1 Природные процессы индукции
- 2 Индуцированные тотипотентные клетки (иТК)
-
3 ИПСК как результат радикального омоложения
- 3.1 Алгоритм для предсказания набора транскрипционных факторов необходимых для преобразования клеток
- 3.2 Стратегии получения ИПСК для клинических испытаний
- 3.3 Система индуцируемого апоптоза для безопасности
- 3.4 Устойчивость к онкогенезу у ИПСК Голого землекопа
- 3.5 Эффективность перепрограммирования в ИПСК
- 3.6 Дифференцировка ИПСК в зрелые клетки в условиях in vivo
- 3.7 Получение клеток хрусталика и сетчатки глаза из ИПСК
- 3.8 Получение из ИПСК легочных эпителиальных клеток
- 3.9 Получение нервных стволовых клеток человека из ИПСК
- 3.10 Получение стволовых клеток почки из ИПСК
- 3.11 Получение остеобластов из ИПСК
- 4 Наивные плюрипотентные стволовые клетки (нПСК)
- 5 Регион-селективные плюрипотентные стволовые клетки (рсПСК)
- 6 Клетки F класса
-
7 Индуцированные прогениторные стволовые клетки
- 7.1 Методы прямой трансдифференцировки
- 7.2 Перепрограммирование путём поэтапного моделирования процессов регенерации
- 7.3 Получение ИПСК и трансдифференцировка с помощью антител
- 7.4 Перепрограммирование бактериями
- 7.5 Условно перепрограммированные клетки (УПК)
- 7.6 Косвенное перепрограммирование клеток (ILC)
- 7.7 Перепрограммирование воздействием на гликопротеин наружней мембраны
- 7.8 Индуцированные стрессом стволовые клетки (ИССК)
- 7.9 Перепрограммирование индуцированное физическим воздействием
- 7.10 Индуцированные нервные стволовые клетки (иНСК)
- 7.11 Прогениторные клетки олигодендроцитов (ПКОД)
- 7.12 Индуцированные кардиомиоциты (иКМ)
- 7.13 Омоложение мышечных стволовых клеток
- 7.14 Индуцированные гепатоциты
- 7.15 Клетки продуцирующие инсулин
- 7.16 Биоинженерия клеток кровеносных сосудов
- 7.17 Прямое перепрограммирование клеток взрослого организма в прогениторные нефроны (ИПН)
- 7.18 Биоинженерия стволовых клеток крови
- 7.19 Иммунные клетки
- 7.20 Омоложение эпителиальных клеток тимуса
- 7.21 Индуцированные стволовые/стромальные клетки мезенхимы (ИМСК)
- 7.22 Индуцированные хондрогенные клетки (ИХОНК)
- 8 Источники соматических клеток
- 9 Культуральная среда для плюрипотентных стволовых клеток свободная от питающих клеток и сыворотки
- 10 Способы доставки репрограммирующих факторов в ядро
- 11 Химически индуцированные плюрипотентные клетки (ХИПСК)
- 12 Краситель для выявления плюрипотентных клеток человека
- 13 Перспективы изучения индуцированных стволовых клеток для медицины
- 14 См. также
- 15 Примечания
- 16 Литература
- 17 Ссылки
Природные процессы индукции
Ещё в 1895 году Томас Морган, удалив один из двух бластомеров лягушки, обнаружил, что оставшаяся часть эмбриона способна, тем не менее, воссоздать цельный эмбрион. Это означало, что клетки, при необходимости, способны изменять направление своей специализации и такое изменение скоординировано. Позднее в 1924 году, Спеманн и Мангольд (Spemann and Mangold) показали, что важнейшую ключевую роль в процессах развития животных играют межклеточные взаимодействия называемые индукцией. Метаплазией называют обратимую замену одного дифференцированного типа клеток на другой тип зрелых дифференцированных клеток. Этот переход от одного типа клеток к другому может быть частью нормального процесса созревания или вызван каким-то индуцирующим его стимулом. Примерами этого перехода можно назвать трансформацию клеток радужной оболочки глаза в линзу в процессе созревания и превращение клеток пигментного эпителия сетчатки в нейральную сетчатку при регенерации глаза у взрослых тритонов. Этот процесс позволяет организму заменить исходные клетки, не подходящие к новым условиям, на новые которые больше подходят к новым условиям. В опытах на клетках имагинальных дисков дрозофилы было обнаружено, что существует ограниченное число стандартных дискретных состояний дифференцировки и клеткам приходится выбирать одно из них. Тот факт, что трансдетерминация (смена пути дифференцировки) часто происходит не в одной, а сразу в группе клеток доказывает, что она вызвана не мутацией, а именно индуцирована.
К настоящему времени удалось выявить минимальные условия и факторы, наличия которых достаточно для индукции каскада молекулярных и клеточных процессов, направляющих дифференцировку и самоорганизацию плюрипотентных клеток в эмбрион. Роль морфогенов, как оказалось, выполняют противоположно направленные градиенты концентрации морфогенетического белка костной ткани (BMP) и белка Nodal.
Некоторые типы зрелых, специализированных клеток взрослого организма способны естественным путём вернуться к стадии стволовой клетки. Например, дифференцированные клетки желудка, называющиеся аделоморфными или «главными клетками» и синтезирующие маркер стволовых клеток Troy, обычно производят пищеварительные жидкости. Однако, они, могут при необходимости превратиться обратно в стволовые клетки, для «ремонтных работ» в случае травм желудка, таких как порез или повреждение вызванное инфекцией. Более того, они осуществляют этот переход даже в отсутствие заметных травм и способны восполнить пул всех клеточных линий желудочного эпителия, по существу, выступая в качестве покоящихся «резервных» стволовых клеток. При повреждении трахеи, дифференцированные эпителиальные клетки дыхательных путей могут вернуться к фенотипу стабильных и функциональных стволовых клеток, если однако, они не имеют непосредственного контакта с базальной стволовой клеткой, которая предотвращает подобную дедифференцировку. Зрелые терминально дифференцированные эпителиальные клетки почки, после травмы, способны дедифференцироваться в свои более ранние версии, а затем снова дифференцироваться в типы клеток нуждавшихся в замене в поврежденной ткани. Макрофаги могут самообновляться путём локальной пролиферации зрелых дифференцированных клеток. Это происходит, когда понижаются концентрации или происходит ингибирование двух факторов транскрипции MafB и c-Maf, препятствующих активации программы самообновления. У тритонов мышечная ткань восстанавливается из специализированных мышечных клеток, которые для этого дедифференцируются забыв свою прежнюю специализацию. Эта способность к регенерации тканей не уменьшается с возрастом, что вероятно связано со способностью тритонов при необходимости образовывать из мышечных клеток новые стволовые клетки.
Следует отметить, что в организме существует небольшой процент стволовых клеток, способных генерировать множество различных типов клеток. Например, мультилинейно-дифференцирующиеся устойчивые к стрессу (англ. muse cell) стволовые клетки взрослого человека обладают способностью к самообновлению и образуют в суспензионной культуре характерные скопления (кластеры) плюрипотентных клеток, которые могут дифференцироваться как in vitro, так и in vivo в энтодермальные, эктодермальные и мезодермальные клетки. Они также легко перепрограммируются в ИПСК.
Подробное описание некоторых других хорошо документированных примеров трансдифференцировки in vivo и их роль в развитии и регенерации рассматриваются в обзоре.
Индуцированные тотипотентные клетки (иТК)
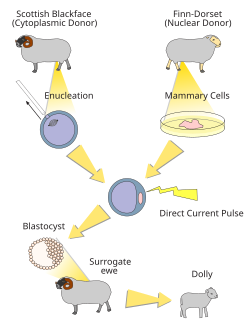
Перепрограммирование в иТК с помощью SCNT
Индуцированные тотипотентные клетки обычно используют для клонирования и получения генетически модифицированных животных. Эти клетки можно получить с помощью перепрограммирования соматических клеток путём переноса ядер соматических клеток (Somatic cell nuclear transfer — SCNT) в ооциты-реципиенты. При этом ооциты не обязательно должны принадлежать тому же виду. Иногда удается использовать ооциты других видов, например овец или поросят. И хотя эффективность межвидовой SCNT была примерно в три раза ниже обычной, такие эмбрионы удавалось довести до стадии бластоцисты. Эффективность перепрограммирования можно повысить в два раза, если за сутки до пересадки остановить мейоз ооцитов-реципиентов с помощью бутиролактона1 в комбинации с нейротрофическим фактором мозга (BDNF). Кроме того, эффективность клонирования может быть значительно повышена, а процедура SCNT упрощена благодаря использованию ингибиторов гистондеацетилазы, таких как трихостатин А и ингибиторов полимеризации цитоскелетного актина таких как цитохалазин В или латранкулин A (latrunculin A). Для полноценного развития образующихся эмбрионов необходимо также предварительно уменьшить метилирование лизина 4 в молекуле гистона H3 в клетках донорах ядра. Кроме того для полноценного развития эмбриона необходимы также экзосомы выделяемые его клетками.
Повторное клонирование на протяжении 25 поколений жизнеспособных мышей с помощью метода SCNT, основанном на добавлении в среду клеточной культуры ингибитора деацетилазы гистонов — трихостатина А, показало, что можно достаточно долго (на протяжении 16 лет) неоднократно повторно клонировать животных без видимого накопления нарушений в геноме.
До настоящего времени бытует представление о возможности преждевременного старения клонированных животных, полученных методом SCNT. Показано, что теломеры у эмбрионов клонированных свиней, полученных с помощью стандартных методов SCNT хуже восстановлены, по сравнению с эмбрионами образующимися по естественному пути. Обработка же трихостатином А значительно увеличивает длину теломер у клонированных свиней и это может быть одним из механизмов, лежащих в основе улучшенного развития клонированных животных, после обработки трихостатином А.
При использовании технологии SCNT разработанной Миталиповым можно получать ЭСК человека используя ядра из фибробластов кожи даже пожилых людей, что открывает широкие перспективы для технологий регенеративной медицины
Разработан метод, открывающий новые возможности для создания генетически модифицированных животных с помощью гаплоидных эмбриональных стволовых клеток, которые могут быть использованы вместо спермы. Для этого из ооцита удаляют ядро. Затем в него вводят микроинъекцией сперму. Из образующейся в результате этого бластоцисты получают гаплоидные эмбриональные стволовые клетки. Эти клетки, синхронизованные в М фазе, вводят в ооцит вместо спермы, в результате чего развивается жизнеспособное потомство. Эти разработки, вместе с данными о возможности неограниченного получения ооцитов из митотически активных половых стволовых клеток, открывают возможность промышленного производства трансгенных сельскохозяйственных животных. Так, в Китае с помощью упрощенной техники клонирования получены трансгенные овцы, у которых улучшено качество мяса и молока за счет увеличения в них незаменимых ненасыщенных жирных кислот, которые снижают риск развития ишемической болезни сердца и необходимы для поддержки глаз и головного мозга. Ген, вызывающий синтез ω-3 полиненасыщенных жирных кислот успешно удалось передать трансгенной овце. Клонирование животных для исследовательских целей в Китае уже приобрело промышленные масштабы. Одних только различных клонов поросят производится порядка 500.
Подобные технологии могут также найти клиническое применения для преодоления цитоплазматических дефектов в ооцитах человека. Например, разработаны технологии, которые могут воспрепятствовать нежелательному наследованию митохондриального заболевания, которое передается следующему поколению. Митохондрии, которые часто называют «электростанцией клетки», содержат генетический материал, который передается от матери к ребёнку. Мутации митохондриальной ДНК могут вызвать диабет, глухоту, заболевания глаз, желудочно-кишечные расстройства, болезни сердца, деменцию и ряд других неврологических заболеваний. Пересадкой ядра из яйцеклетки одного человека (несущей дефектную митохондриальную ДНК) в другую (здоровую) можно эффективно заменить цитоплазму клетки и вместе с ней митохондрии (и их ДНК). Полученная таким образом яйцеклетка может рассматриваться как имеющая двух матерей. Эмбрион образующийся после оплодотворения такой яйцеклетки будет иметь здоровую митохондриальную ДНК. Однако насколько оправданы подобные манипуляции с клетками человека с точки зрения биоэтики пока не ясно.
Подробнее о новейших достижениях техники клонирования и получении тотипотентных клеток с помощью SCNT см.:
Перепрограммирование в иТК без помощи SCNT
До недавнего времени получить тотипотентные клетки удавалось лишь с помощью SCNT. Однако появились работы где было продемонстрировано получение иТК с помощью перепрограммирования факторами Яманаки in vivo, а также in vitro с помощью таких эпигенетических факторов ооцита как зародышевая изоформа гистонов. Перевести эмбриональные стволовые клетки в состояние тотипотентности характерное для ранних эмбрионов 2-клеточной стадии можно также путём подавления активности CAF-1 необходимой для сборки хроматина. От превращения ЭСК и ИПСК в тотипотентные клетки, способные дать начало таким внеэмбриональным тканям как плацента и желточный мешок, плюрипотентные стволовые клетки, по-видимому, так же удерживает микроРНК-34а. Ингибирование её синтеза приводит к активации экспрессии эндогенного ретровируса MuERV-L и расширяет потенциал плюрипотентных стволовых клеток до способностей клеток стадии двух бластомеров.
Разработан химический коктейль для получения так называемых плюрипотентных стволовых клеток с расширенными возможностями по дифференцировке — они могут дать начало как клеткам эмбриона, так и внеэмбриональным тканям. Такие супертотипотентные клетки (иСТК) могут быть использованы для получения химер с целью выращивания органов в организме животных
Получение репродуктивных клеток из ИПСК
Используя среды, содержащие ретиноевую кислоту и фолликулярную жидкость свиньи, можно получить in vitro, дифференцировкой из ИПСК, клетки ранних стадий гаметогенеза подобные репродуктивным клеткам, из которых образуются сперма и ооциты. Примечательно, что для образования примордиальных половых клеток человека требуется активность двух ключевых регуляторов: гена SOX17 направляющего дифференцировку в сторону образования предшественников половых клеток и Blimp1 подавляющего энтодермальные и другие соматические гены во время этой специализации.
В статье китайских ученых с первым автором Чжоу (Zhou) описана технология дифференциации мышиных эмбриональных стволовых клеток которые претерпевают мейоз in vitro превращаясь в гаплоидные сперматиды, способные к оплодотворению, о чём свидетельствовало получение с их помощью жизнеспособного и фертильного потомства.
Подробный обзор методов искусственного получения мужских половых клеток можно найти в статье Hou с соавт. и Irie, Kim, Surani
Разработана технология позволяющая получать зрелые ооциты in vitro из эмбриональных стволовых клеток, а также из индуцированных плюрипотентных стволовых клеток, полученных из взрослых фибробластов взятых с кончика мышиного хвоста. Более того оплодотворив такие яйцеклетки in vitro и подсадив их в матку мыши удалось с выходом в 1 % получить жизнеспособное потомство. Эта технология послужит платформой для выяснения молекулярных механизмов, лежащих в основе тотипотентности и для разработки методов производства ооцитов из других (в том числе редких) видов млекопитающих в лабораторных условиях.
ИПСК как результат радикального омоложения

Впервые ИПСК были получены в виде перевиваемой тератокарциномы, индуцированной трансплантатом, взятым из мышиных эмбрионов. Было доказано, что тератокарциномы образуются из соматических клеток. Тот факт, что из клеток тератокарциномы можно получить нормальную мышь доказывал их плюрипотентность. Оказалось, что клетки тератокарциномы, выделяя в культуральную среду различные факторы, способны поддерживать культуру плюрипотентных стволовых клеток эмбриона в недифференцированном состоянии. Таким образом, ещё в 1980-е годы стало ясно, что трансплантация плюрипотентных или эмбриональных стволовых клеток во взрослый организм млекопитающих обычно приводит к образованию тератомы, которая затем может превратиться в злокачественную опухоль — тератокарциному. Если, однако, поместить клетки тератокарциномы в ранний зародыш млекопитающего (на стадии бластоцисты), то они включаются в состав клеточной массы бластоцисты и из такого химерного (то есть состоящего из клеток от разных организмов) эмбриона нередко развивается нормальное химерное животное. Почти во всех органах и тканях которого часть дифференцированных клеток происходит из клеток тератокарциномы, которые совместно с клетками нормального происхождения участвуют в построении здорового организма. Это свидетельствовало о том, что причиной образования тератомы является диссонанс в стадии развития донорных клеток и окружающих их клеток реципиента (так называемой ниши). Уже тогда, используя ретровирусные векторы, удалось ввести инородные гены в мышиные химеры, полученные с помощью клеток тератокарциномы.

В августе 2006 года японские исследователи сумели превратить клетки мышиной кожи (фибробласты) в индуцированные плюрипотентные стволовые клетки — ИПСК (induced pluripotent stem cells — iPSC), используя для модификации клетки всего четыре репрограммирующих фактора: Oct4, Klf4, Sox2 и c-Myc, доставленных в ядро ретровирусами. Этим они доказали, что гиперэкспрессия небольшого количества факторов иногда может подтолкнуть клетки к переходу в новое стабильное состояние, связанному с изменениями активности тысяч генов. По своим свойствам ИПСК оказались очень похожи на эмбриональные стволовые клетки (ЭСК). Так, сравнение протеома и фосфопротеома ЭСК и ИПСК, проведенное на 4-х линиях человеческих эмбриональных стволовых клеток и 4-х линиях индуцированных плюрипотентных стволовых клеток, показало, что большинство идентифицированных белков и участков фосфорилирования в белках всех линий совпадают. Хотя были и небольшие, но статистически воспроизводимые различия, свидетельствующие об определённом функциональном различии. Не было отмечено и особых изменений в последовательности ДНК, особенно если ИПСК были получены с помощью неинтегрирующихся в геном плазмид. Позднее, с развитием технологии перепрограммирования, лучшим доказательством идентичности ИПСК и ЭСК стала возможность получения взрослой мыши полностью из некоторых линий ИПСК. Несмотря на то, что рядом исследований была доказана идентичность ЭСК и ИПСК, получаемые клоны сильно отличаются друг от друга и не для всех из них можно доказать идентичность с ЭСК, далеко не все клоны способны дать жизнь химерным мышам или подвергнуться эффективной дифференциации в те или иные соматические клетки. Одной из причин таких различий является разница между составом транскрипционных факторов при репрограммировании в ИПСК и набором факторов в материнском ооците. К числу таких «упущенных» факторов относится, в частности способствующий процессу репрограммирования особый, характерный для ооцитов линкерный (связующий, компонующий нуклеосомы) гистон H1foo. Замена одного из факторов Яманаки, а именно c-Myc на H1foo, значительно повысили количество и качество получаемых клонов ИПСК — они стали более однородными по свойствам, из них чаще стали получаться мыши-химеры.
Команда, возглавляемая Гансом Шелером (который открыл ген Oct4 в 1989 году), показала, что избыточная экспрессия Oct4 во время перепрограммирования ухудшает качество ИПСК. По сравнению с OSKM (Oct4, Sox2, Klf4 и c-Myc), которые показывают патологические модели импринтинга и дифференцировки, перепрограммирование SKM (Sox2, Klf4 и c-Myc) генерирует ИПСК с высоким потенциалом развития (почти в 20 раз выше, чем у OSKM) эквивалентно эмбриональным стволовым клеткам, что доказано по их способности генерировать мышей полностью состоящих из ИПСК посредством метода тетраплоидной комплементации эмбрионов.
Важным преимуществом ИПСК перед ЭСК является то, что они могут быть получены из клеток взрослого организма, а не из эмбриона. Поэтому стало возможным получать ИПСК от взрослых и даже пожилых пациентов. Перепрограммирование соматических клеток в ИПСК приводит к их омоложению о чём свидетельствуют данные исследования теломеров— концевых участков хромосом состоящих из коротких следующих друг за другом повторов эволюционно консервативной последовательности ДНК. Выяснилось, что перепрограммирование приводит к удлинению теломеров и их нормальному укорочению по мере дифференцировки ИПСК обратно в фибробласты. Таким образом, при индуцированной плюрипотенции восстанавливается эмбриональная длина теломеров, а значит, увеличивается потенциальное число делений клетки, ограниченное так называемым лимитом Хайфлика (Hayflick limit). Более того омолаживаются и митохондрии клетки при этом восстанавливается характерный для молодых клеток уровень дыхания Поэтому технологию получения ИПСК следует рассматривать как способ радикального омоложения. Из-за диссонанса в стадии развития омоложенных клеток и окружающих их старых клеток реципиента, инъекция пациенту его же собственных ИПСК, обычно приводит к иммунной реакции, что может быть использовано в медицинских целях, или образованию опухолей типа тератомы. Одной из причин иммуногенности аутологичных ИПСК и ЭСК считается группа из 9 генов (Hormad1, Zg16, Cyp3a11, Lce1f, Spt1, Lce3a,Chi3L4, Olr1, Retn), синтез которых повышен в тератомах, полученных из этих клеток Очевидно, некоторые клетки, дифференцированные из ИПСК и ЭСК, продолжают синтезировать эмбриональные изоформы белков и неадекватно интерпретируют сигналы окружающих их клеток реципиента. В связи c этим следует отметить, что образование тератомы из плюрипотентных стволовых клеток может быть вызвано низкой активностью фермента PTEN, способствующей выживанию, в процессе дифференцировки, небольшой популяции (не превышающей 0,1-5 % от общей численности клеток) высоко онкогенных клеток карциномы, инициирующих тератомы. Выживание этих инициирующих тератомы клеток связано с недостаточной репрессией Nanog, а также с повышением метаболизма глюкозы и холестерина. Эти, инициирующие образование тератом, клетки характеризуются также более низким соотношением p53/p21 по сравнению с неонкогенными клетками.
Недавно методом отбора удалось найти небольшие молекулы (цитотоксические селективные ингибиторы плюрипотентных стволовых клеток человека), которые предотвращают образование тератомы у мышей после трансплантации им плюрипотентных стволовых клеток человека. Самое мощное и селективное из этих соединений — PluriSIn #1, вызывало ингибирование стеароил-КоА десатуразы (ключевого фермента в биосинтезе олеиновой кислоты), что в конечном итоге приводило к апоптозу плюрипотентных стволовых клеток. С помощью этой молекулы удается выборочно удалить из культуры недифференцированные клетки.. Ещё одной молекулой избирательно удаляющей недифференцированные клетки является STF-31, являющийся ингибитором GLUT1. Эффективной стратегией избирательного устранения плюрипотентных клеток, которые способны дать начало тератоме является ингибирование характерных для этих клеток антиапоптотических факторов, таких как сурвивин или Bcl10. Обработкой малыми молекулами, которые могут ингибировать эти антиапоптотические факторы, можно добиться селективного удаления подобных клеток вызвав их апоптоз. В частности, одной обработки смешанной популяции химическими ингибиторами сурвивина (такими как, например, кверцетин или YM155) достаточно чтобы вызвать избирательную и полную гибель недифференцированных клеток, вызванную накоплением р53 в митохондриах. Этого, по мнению авторов исследования, достаточно, чтобы предотвратить образование тератомы после трансплантации клеток полученных из ИПСК. Тем не менее, маловероятно, что какая либо, пусть даже самая изощренная, предварительная очистка, способна обезопасить подсадку ИПСК или ЭСК, так как при избирательном удалении плюрипотентных клеток, они вновь довольно быстро возникают путём превращения дифференцированных клеток обратно в стволовые (к обратному переходу может в частности подтолкнуть гипоксия), что приводит к образованию опухоли. Это может быть связано с нарушением регуляции осуществляемой микро РНК let-7 по отношению к её мишени — белку Nr6a1 (известному также как ядерный фактор зародышевых клеток — GCNF), являющемуся эмбриональным репрессором транскрипции генов плюрипотентности, который необходим для правильной дифференцировки индуцированных плюрипотентных клеток. Обнаружена также малая молекула названная Дисплюригеном (Displurigen), которая воздействуя на белок теплового шока HSPA8 (Heat shock 70 kDa protein 8), необходимый для связывания OCT4 с ДНК, способна вывести клетку из состояния плюрипотентности. Ещё один способ предотвратить образование тератомы — это вызвать в пересаживаемой клетке ИПСК гиперэкспрессию гена CREG
Использование ИПСК для клеточной терапии пока ограничено. Тем не менее, они могут быть использованы для целого ряда иных целей — включая моделирование болезней, скрининг (селективный отбор) лекарств, проверку токсичности различных препаратов. Важными факторами для получения высококачественных ИПСК являются определённые небольшие молекулы, способствующие сохранению геномной целостности, образующихся при перепрограммировании ИПСК, путём ингибирования двухцепочечных разрывов ДНК и активации гена Zscan4, содействующего процессам репарации ДНК. Перепрограммирование вызывает репликативный стресс, который можно снизить повысив уровень чекпоинт киназы 1 (CHK1), благодаря чему повышается качество и эффективность образования ИПСК. Кроме того добавление во время перепрограммирования нуклеозидов позволяет снизить повреждения ДНК и число геномных перестроек в получаемых ИПСК
Интересно отметить, что ткани, выращенные из ИПСК, помещенных в «химерные» эмбрионы на ранних стадиях развития мыши, практически не вызывают иммунного ответа (после того, как эмбрионы выросли во взрослых мышей) и пригодны для аутологичной трансплантации, причем даже в том случае когда ИПСК получены от очень старых животных. В то же время, полное перепрограммирование взрослых клеток в тканях у мышей in vivo путём временной активации факторов Oct4, Sox2, Klf4 и с-Мус, приводит к образованию в различных органах множества тератом. Смотри рисунок. Более того, частичное перепрограммирование клеток в ИПСК in vivo показало, что неполное перепрограммирование приводит к эпигенетическим изменениям (нарушению репрессии Поликомб целей и изменению метилирования ДНК) в клетках, которые ведут к развитию рака

Алгоритм для предсказания набора транскрипционных факторов необходимых для преобразования клеток
Определение уникального набора транскрипционных факторов, которые необходимы для репрограммирования клеток представляет собой длительный и дорогостоящий процесс. Международная группа исследователей разработала алгоритм, называемый Магрифи (Mogrify), который помогает предсказать оптимальный набор клеточных факторов, необходимых для преобразования одного типа клеток человека в другой. Появился также алгоритм, который предсказывает не только транскрипционные факторы необходимые для перепрограммирования, но также и идеальный выбор времени для применения этих факторов.
Стратегии получения ИПСК для клинических испытаний
Разработаны критерии качества и стратегия производства ИПСК для клинических испытаний, так называемая cGMP (англ. current Good Manufacturing Practice).
Система индуцируемого апоптоза для безопасности
Чтобы обезопасить применение ИПСК в клинике, было предложено одновременно с перепрограммированием клеток пациента в ИПСК, вводить в эти клетки индуцируемый малой молекулой ген каспазы-9 (IC9) для запуска каскадов апоптоза для самоубийства клеток образованных из этих ИПСК. Такой «предохранитель» позволит избавляться от омоложенных клеток после того как они выполнили свою терапевтическую функцию или в случае образования опухоли из этих клеток.
Устойчивость к онкогенезу у ИПСК Голого землекопа
У голых землекопов уровень заболеваемости раком крайне низок по сравнению с другими млекопитающими. Обнаружено, что у ИПСК этого животного ослаблена способность к образованию тератом при трансплантации, что может быть связано:
- с видоспецифической активацией супрессора опухоли ARF (англ. alternative reading frame), который является продуктом альтернативной рамки считывания гена CDKN2A (другой продукт этого гена — маркер старения белок p16), а также
- с мутацией, приводящей к разрушению онкогена ERAS, являющегося аналогом Ras и отвечающего за онкогенность ЭСК.
Более того, удалось найти сигнальный путь ASIS (англ. ARF suppression-induced senescence), с помощью которого вероятно удастся защитить ИПСК от возникновения из них опухолей
Эффективность перепрограммирования в ИПСК
До настоящего времени недостаточно понятно, почему эффективность перепрограммирования с помощью факторов транскрипции значительно ниже, чем при пересадке ядра в ооцит. Показано, что большинство фибробластов кожи взрослого человека начинают процесс перепрограммирования сразу после обработки трансгенами Яманаки (Oct4, Sox2, Klf4, и c-Myc). Тем не менее, только небольшая часть (~ 1 %) из этих «новоиспеченных» ИПСК образуют впоследствии колонии ИПСК. Причиной, понижающего эффективность перепрограммирования, возврата большинства клеток к состоянию дифференцировки может быть:
- недостаточная деятельность активируемой цитидиндезаминазы (AID) из-за чего клетки не могут стабилизироваться и долго поддерживать состояние плюрипотенции.
- недостаточная активность гена SMC1 кодирующего один из белков когезина (необходимого для образования внутрихромосомной петли сближающей промоутер гена с последующим энхансером, что необходимо для активации эндогенных генов плюрипотентности), делает невозможным достижение плюрипотентности
- важную роль на поздних этапах перепрограммирования играют и ферментативные модификации гистонов. Показано, что необходимым условием эффективного перепрограммирования является подавление переносчика гистонов CAF-1 и белкового комплекса ремоделирования нуклеосом и деацетилирования (nucleosome remodeling and deacetylation — NuRD. Избыточная экспрессия субъединицы NuRD, называемой Mbd3, ингибирует индукцию ИПСК. Причиной этого является деацетилирование комплексом NuRD лизина 27 в молекуле гистона Н3К27ac, что позволяет Поликомб Репрессорному комплексу 2 (PRC2) осуществить триметилирование лизина 27 в гистоне H3, приводящее, в конечном счете, к ингибированию ряда генов-маркеров плюрипотентности , в том числе генов Oct4 и Nanog. Ингибирование Mbd3, с другой стороны, повышает эффективность перепрограммирования и способствует образованию плюрипотентных стволовых клеток, которые способны генерировать жизнеспособных химерных мышей, даже в случае отсутствия с-Мус или Sox2. Очевидно, Mbd3/NuRD исполняет роль эпигенетического регулятора, который ограничивает экспрессию ключевых генов плюрипотентности. Поэтому подавление Mbd3/NuRD (например, с помощью бутирата, вальпроевой кислоты, субероиланилидгидроксамовой кислоты или трихостатина А) может стать мощным средством для повышения эффективности и точности перепрограммирования. Действительно, подавив Mbd3 удалось впервые осуществить детерминированное и синхронизированное перепрограммирование клеток кожи мыши и человека в ИПСК в течение всего семи дней и с невиданной ранее эффективностью — около 100 %
Найден фактор BRD3 (bromodomain-containing protein 3), который опознаёт «коды» ацетилированных гистонов в хромосоме, а также активирует большой набор митотических генов, повышая таким образом митотическую активность клетки. Этот фактор позволил более чем в 20 раз повысить эффективность выхода ИПСК, сократить длительность перепрограммирования до нескольких дней и повысить качество перепрограммирования. Как отмечено выше повысить эффективность репрограммирования позволяет также замена с-Мус на H1foo. В случае когда требуется репрограммировать клетки пожилых пациентов повысить эффективность позволяет ингибирование H3K79 гистон метилтрансферазы называемой DOT1L (Disruptor of telomeric silencing 1-like)
Элитные клетки
В первичных после биопсии культурах клеток при перепрограммировании лишь очень немногие клетки способны превратиться в ИПСК, и тех из них, которые такой способностью обладают, называют «элитными» клетками. Ученые нашли способ получения таких элитных клеток из соматических с помощью фактора C/EBPα (CCAAT/enhancer binding protein-α). В первичной культуре мышиных В-клеток непродолжительная экспрессия C/EBPα с последующим перепрограммированием факторами Яманаки позволила добиться 100-кратного увеличения эффективности перепрограммирования в плюрипотентные клетки, причём с участием 95 % клеточной популяции. Такие искусственно созданные элитные клетки очень похожи на белые кровяные прогениторные клетки-предшественники костного мозга, известные как миелобласты.
Дифференцировка ИПСК в зрелые клетки в условиях in vivo
В тератоме
Тот факт, что ИПСК человека способны к образованию тератом не только в теле человека, но и в организме некоторых животных, в частности в организме мыши или свиньи, позволил разработать метод дифференцировки ИПСК в условиях in vivo. Для этого ИПСК вводят, вместе с клетками индуцирующими направленную дифференцировку, генмодифицированной свинье или мыши, у которой подавлена активация иммунной системы на клетки человека, а затем, вырезав образовавшуюся тератому, выделяют из неё необходимые дифференцированные клетки человека, используя моноклональные антитела к тканеспецифичным маркерам на поверхности полученных клеток. Этот метод был успешно использован для получения функциональных мышечных, а также миелоидных, лимфоидных и эритроидных клеток человека пригодных для трансплантации (пока только мышам). Таким образом, доказана возможность производства in vivo из клеток пациента необходимых ему дифференцированных клеток для трансплантации, изготовления антител или скрининга лекарственных средств. Используя перевиваемую генмодифицированную тератому с гиперэкспрессией факторов Gfi1b, c-Fos и Gata2 можно неоднократно трансплантировать мышкам тератому и на протяжении длительного времени стабильно получать полностью функциональные мышиные гематопоэтические стволовые клетки
Используя лектин rBC2LCN избирательно связывающий ИПСК, или же MitoBloCK-6 и /или PluriSIn #1 можно очистить полученные прогениторные клетки от плюрипотентных клеток образующих тератому. Тот факт, что дифференцировка проходит в условиях тератомы позволяет надеяться, что полученные клетки достаточно устойчивы к стимулам способным запустить их обратный переход к дедифференцированному (плюрипотентному) состоянию, а значит безопасны. Беспокойство, однако, вызывает тот факт, что «воспитанные» в тератоме у животных человеческие клетки за время своего «воспитания», по всей вероятности, поглощают значительное количество экзосом произведенных окружающими клетками организма носителя тератомы, а значит, попав в организм человека, могут повести себя неадекватно.
Методика основанная на обнаружении ген-репортер-GFP-положительных клеток в тератоме, полученной из ИПСК, позволит идентифицировать и вырастить культуры ткани, используя индуцированные взрослые стволовые клетки различных типов, выделение которых ранее было затруднительно.
В организме животных-биоинкубаторов
Весьма перспективной средой для первоначальной дифференцировки ИПСК in vivo могут оказаться куриные эмбрионы. Есть доказательства того, что микросреда этих эмбрионов оказывает анти-онкогенное действие на человеческие клетки и намного лучше чем условия in vitro
Разработана технология «дозревания», полученных из ИПСК в условиях in vitro человеческих прогениторных клеток кардиомиоцитов, путем ксенотрансплантации их в организм новорожденных крыс, используемых в качестве in vivo биоинкубатора. Такое «дозревание» занимает ~6 недель
См. также: Robert Lanza, Michael West (2013) Method for facilitating the production of differentiated cell types and tissues from embryonic and adult pluripotent and multipotent cells. Patent US 20130058900 A1
Получение клеток хрусталика и сетчатки глаза из ИПСК
В ближайшее время предполагается приступить к клиническим испытаниям, призванным продемонстрировать безопасность использования ИПСК для клеточной терапии людей с катарактой а также с возрастной дегенерацией жёлтого пятна — заболевания, которое повреждая сетчатку, может привести к слепоте. Описаны методы получения из ИПСК клеток хрусталика и сетчатки и способы их использования для клеточной терапии, которая по крайней мере на 6 недель улучшала зрение у подопытных животных.
Получение из ИПСК легочных эпителиальных клеток
Хронические заболевания легких, такие как идиопатический фиброзирующий альвеолит, Силикоз, хроническая обструктивная болезнь легких и бронхиальная астма входят в число основных причин инвалидности и смертности. Поэтому исследователи ищут пути эффективной клеточной терапии и тканевой инженерии легких, которые бы позволили бороться с этими заболеваниями. Были разработаны способы получения различных типов легочных клеток из ИПСК, которые могут быть взяты за основу для получения терапевтических клеток из материала, полученного от пациента.
Получение нервных стволовых клеток человека из ИПСК
Юань и коллеги сообщили, что нервные стволовые клетки человека, индуцированные из ИПСК с помощью ретиноевой кислоты в бессывороточной среде имеют стабильный нейронный фенотип. После трансплантации крысам со смоделированным ишемическим инсультом, эти клетки не только выжили, но и мигрировали в зону ишемии мозга, где дифференцировались в зрелые нервные клетки, что оказало благотворное влияние на функциональное восстановление утраченных от повреждения в результате инсульта неврологических функций.
Получение стволовых клеток почки из ИПСК
Разработана система для быстрого (за 3 дня) и эффективного (70 %-80 % популяции) превращения ИПСК в клоны характерные для клеток почки с помощью ингибитора CHIR99021 и некоторых ростовых факторов. Более того, удалось излечивать в опытах на мышах острые поражения почек, используя стволовые клетки почки, полученные из ИПСК.
Получение остеобластов из ИПСК
Известно что аденозин и его рецепторы, в частности A2bR играют важную роль в регенерации костных переломов. Простое добавление в культуральную среду аденозина позволило превратить человеческие ИПСК в остеобласты. При трансплантации этих остеобластов мышке, с использованием макропористой синтетической матрицы, остеобласты полученные из ИПСК, участвовали в регенерации повреждений кости образуя новые ткани и стимулируя кальцификацию. При этом не наблюдалось образования тератом, что очевидно свидетельствует о 100 % дифференцировке клеток ИПСК в остеобласты.
Наивные плюрипотентные стволовые клетки (нПСК)
Человеческие плюрипотентные стволовые клетки, независимо от того, получены ли они из бластоцисты или являются результатом перепрограммирования соматических клеток, существенно отличаются от классических мышиных эмбриональных стволовых клеток и по мнению ряда исследователей представляют более позднюю стадию развития эпибласта. Удалось получить нПСК у которых утеряна эпигенетическая «память» метилирования ДНК как гаметы (ооцита), так и человеческой бластоцисты. Такие клетки в отличие от ИПСК не имеют антиген SSEA4 (Stage Specific Embryonic Antigen 4). Перевести ЭСК и ИПСК человека в наивное состояние позволяет гиперэкспрессия фактора YAP (Yes-associated protein). Гиперэкспрессию YAP с получением наивного состояния можно также имитировать путём добавления к культуральной среде (лизофосфатидной кислоты (LPA), являющейся активатором YAP.
Репрограммирование человеческих ЭСК и ИПСК с помощью рекомбинантного, усеченного человеческого NME7 (найденного в семенниках фактора, содержащего два домена нуклеозид дифосфат киназы (NDPK) и способного связываться с расщепленной формой трансмембранного рецептора MUC1, называемой MUC1*) позволило получить стабильно наивные клетки, которые более пригодны для широкомасштабного клонирования и имеют расширенный потенциал дифференциации. На основе таких клеток можно создать «фабрики клеток» для промышленного производства продукции необходимой для нужд клеточной терапии.
Регион-селективные плюрипотентные стволовые клетки (рсПСК)
Ву и его коллеги обнаружили, что комбинация свободной от сыворотки среды, фактора роста фибробластов 2 (FGF2) и ингибитора сигнальных путей Wnt позволяет получить в результате устойчивую линию рсПСК (регион-селективных плюрипотентных стволовых клеток, по англ rsPSCs) клеток человека. По транскриптому эти клетки напоминали таковые из задних клеток раннего эмбриона мыши. Трансплантация этих клеток в 7,5-дневные эмбрионы мыши привела к их эффективному включению в задний, но не в другие части эмбриона. После 36 часов культивирования этих химерных эмбрионов, клетки рсПСК проявили способность к пролиферации и способность к дифференцировке в ткани трёх зародышевых листков. Хотя исследователи остановили дальнейшую дифференцировку этих клеток, предполагается, что каждый из образованных этими клетками зародышевых листков способен дать начало определённым тканям и органам. Важно отметить, что в отличие от других человеческих стволовых клеток, которые, как правило, не удается интегрировать в эмбрион мыши, человеческие rsPSCs способны к такой интеграции и к развитию в ранние стадии тканей человека.
Клетки F класса
Клетки F класса в отличие от ИПСК не способны включаться в ткани организма и участвовать в построении химерного организма. Тем не менее они удовлетворяют другому тесту на плюрипотентность — способны образовывать тератомы. По сравнению с обычными стволовыми клетками подобными эмбриональным и ИПСК,клетки F-типа растут в лаборатории быстрее и их выращивать проще и дешевле — их можно просто поместить в большой сосуд с питательной средой и вырастить за несколько дней или часов, а не за несколько недель как обычные ИПСК.
Индуцированные прогениторные стволовые клетки
Методы прямой трансдифференцировки
В связи с тем, что использование ИПСК для клеточной терапии сопряжено с большим риском опухолей и рака, необходима разработка методов получения более безопасных клеточных линий, пригодных для применения в клинике. Альтернативой методам ИПСК стала техника так называемого «прямого репрограммирования», то есть индуцируемой определёнными факторами прямой трансдифференцировки, без предварительного прохождения клеток через стадии плюрипотентного состояния. Основу для такого подхода заложили исследования Тейлор и Джонса (Taylor and Jones), показавших, что воздействие 5-азацитидина — реактива, вызывающего деметилирование ДНК — на бессмертную линию клеток мышиных эмбриональных фибробластов способно вызвать образование миогенных, хондрогенных и адипогенных клонов и Вейнтрауба с соавторами, обнаруживших, что для репрограммирования достаточно активации всего одного гена, позднее названного MyoD1. По сравнению с ИПСК, для получения которых требуются не менее двух недель, образование индуцированных прогениторных клеток происходит сравнительно быстро — иногда за несколько дней. Эффективность перепрограммирования также обычно во много раз выше. Для этого перепрограммирования не всегда требуется деление клетки. Но главное, это то, что получаемые в результате перепрограммирования мультипотентные соматические стволовые клетки более пригодны для клеточной терапии, так как не образуют тератомы. См. также обзор
Трансдифференцировка с помощью 5-азацитидина и тромбоцитарного фактора роста
Разработан метод получения, так называемых, индуцированных мультипотентных стволовых клеток (ИМПСК) путём непродолжительной обработки постнатальных стволовых клеток костного мозга и жировых клеток комбинацией фактора роста (тромбоцитарный фактор роста — АВ (PDGF-AB)) и 5-азацитидина. Авторы этого исследования утверждают, что: «В отличие от первичных мезенхимальных стволовых клеток, которые хотя и используются в клинической практике для содействия восстановлению тканей, но не способны сами включаться в эту ткань, ИМПСК способны к непосредственному участию процессах регенерации тканей и при этом не образуют опухолей», в связи с чем «могут быть использованы для регенерации различных тканей»
Трансдифференцировка зрелых клеток всего одним фактором транскрипции
Особенностью нематоды Caenorhabditis elegans является настолько жёсткая программа развития, что соматическая клетка, находящаяся в определённом участке организма, как правило, имеет одинаковую родословную у всех особей. При этом зрелые клетки, в отличие от ранних эмбриональных клеток, обычно очень устойчивы к изменению их фенотипа. Тем не менее, обнаружено, что как у интактных личинок, так и у неповреждённых взрослых нематод краткосрочный синтез всего одного фактора транскрипции, а именно фактора ELT-7 GATA может превратить фенотип полностью дифференцированной, высокоспециализированной не-энтодермальной клетки фаринкса (глотки) в фенотип полностью дифференцированной энтодермальной клетки кишечника. Это превращение происходит «в одну стадию» — путём прямой трансдифференцировки, без каких-либо промежуточных стадий дедифференцировки.
Трансдифференцировка с помощью CRISPR-опосредованного активатора
Фенотип клетки можно изменять с помощью редактирования эпигенома. Например, путём активации определённых эндогенных генов, используя для этого CRISPR — опосредованный активатор. Если соединить домен dCas9 (который изменён таким образом, что он больше не режет ДНК, но все ещё может находить и связываться с конкретными последовательностями ДНК) с активатором транскрипции (таким как p65HSF1), то можно с большой точностью изменять эндогенную экспрессию конкретных генов. Например, активируя только один эндогенный ген Sox2 или Oct4 удалось получить из фибробластов мыши ИПСК с выходом в 0,1 %. Пользуясь подобным методом, Вэй и др. усиливали экспрессию эндогенных генов Cdx2 и Gata6, воздействуя на них с помощью CRISPR-опосредованных активаторов, и таким образом, сумели осуществить прямое репрограммирование мышиных эмбриональных стволовых клеток в две внезародышевые линии, а именно в типичные трофобласты и клетки внеэмбриональной энтодермы. Аналогичным образом активация эндогенных генов Brn2, Ascl1, и Myt1l позволила преобразовать эмбриональные фибробласты в индуцированные нервные клетки.
Перепрограммирование путём поэтапного моделирования процессов регенерации
Ещё один способ перепрограммирования заключается в поэтапном моделировании на скелетной мышце млекопитающих процессов, которые происходят у амфибий при регенерации конечности. Таким образом, с помощью химических веществ: миосеверина (myoseverin), реверсина (2-(4-морфолиноанилино)-6-циклогексиламинопурина) и некоторых других веществ, в условиях культуры мышечных клеток млекопитающих, которые, как известно, не способны регенерировать конечности, удалось индуцировать процессы аналогичные тем которые протекают при регенерации конечностей у амфибий и получить предшественники мышечных, костных, жировых и нервных клеток.
Получение ИПСК и трансдифференцировка с помощью антител
Обнаружены моноклональные антитела, способные преобразовывать стволовые клетки костного мозга непосредственно в прогениторные клетки нейронов головного мозга.
Для этой трансдифференцировки как оказалось достаточно всего одного белка — антитела имитирующего фактор GCSF. Для поиска подобных антител используется специальный метод селекции антител.
Идентифицированы антитела, которые могут во время перепрограммирования эмбриональных фибробластов мыши в ИПСК соответственно заменить три из четырёх факторов перепрограммирования Sox2, c-Myc или Oct4. Найти замену четвёртому фактору Klf4 пока не удалось. Более того Sox2-замещающее антитело действуя как антагонист к связанному с мембраной белку Basp1, тем самым активирует подавленные им ядерные факторы WT1, Esrrb и Lin28a (Lin28) независимо от Sox2.
Перепрограммирование бактериями
Желудочно-кишечный тракт человека колонизирован обширным сообществом бактерий симбионтов и комменсалов. Исследователи продемонстрировали феномен перепрограммирования соматических клеток бактериями и выработку мультипотентных клеток из клеток кожи человека, под воздействием молочнокислых бактерий. Выяснилось, что подобная клеточная трансдифференцировка вызвана рибосомами и «может произойти под воздействием бактерий, которые проглатываются и перевариваются клетками-хозяевами, что приводит к стрессу от попадания чужеродных рибосом и стимулирует клеточную пластичность».
Условно перепрограммированные клетки (УПК)
Ричард Шлегель и его исследовательская группа разработали метод, который позволяет размножать in vitro культуру клеток похожих на взрослые стволовые клетки, без каких-либо генетических манипуляций. Они показали, что под воздействием облученных фибробластов (см. обзоры и) и ингибитора Rho киназы — Y-27632, первичные эпителиальные клетки млекопитающих переходят к состоянию неограниченной пролиферации (что, по мнению авторов, связано с ростом концентрации β-катенина в ядре и снижением Notch сигнализации). Индукция УПК происходит довольно быстро (в течение 2 дней) и является результатом «перепрограммирования» всей клеточной популяции, а не одной из её субпопуляций. При этом в УПК не наблюдалась характерная для ИПСК или эмбриональных стволовых клеток (ЭСК) активация синтеза Sox2, Oct4, Nanog, и Klf4. Эта индукция УПК обратима — достаточно удалить Y-27632 и облученные фибробласты, чтобы клетки перешли к обычной дифференцировке. Обнаружено, что факторы, вызывающие индукцию условно перепрограммированных клеток, переходят из «питающих» клеток подложки в культуральную среду в результате обусловленного радиацией апоптоза этих клеток. Этот метод может иметь большое будущее в регенеративной медицине, так как эти клетки в отличие от ИПСК не образуют опухоли. Так, например, используя технологию условно-перепрограммированных клеток, исследователи смогли найти эффективную терапию для пациента с редким типом опухоли легких.
Иной подход к получению условно перепрограммированных клеток заключается в ингибировании мембранного белка CD47, являющегося рецептором тромбоспондина-1. Показано, что потеря CD47 снимает запрет на устойчивую пролиферацию первичных мышиных эндотелиальных клеток, повышая частоту их асимметричного деления, а также позволяет этим клеткам спонтанно перепрограммироваться в мультипотентные клетки формирующие эмбриональные тела. Нокдаун гена CD47 резко увеличивает в клетках уровни мРНК с-Мус и других факторов перепрограммирования Яманаки как in vitro, так и in vivo. Очевидно, тромбоспондин-1 является ключевым сигналом нишы, который подавляет способность стволовых клеток к самообновлению влияя на них через CD47. Поэтому антагонисты CD47 могут активировать самообновление и перепрограммирование клеток выключая механизмы негативной регуляции с-Мус и других факторов транскрипции стволовых клеток По мнению авторов исследования, образующиеся при этом мультипотентные клетки не образуют тератом.
Интересно отметить, что in vivo блокада CD47 с помощью антисмыслового морфолино повышает выживаемость мышей, тело которых подверглось воздействию летальной дозы облучения. Эта устойчивость к радиации обусловлена увеличением пролиферативной способности клеток крови, образующихся из костного мозга, и активации защитной аутофагии радиочувствительных желудочно-кишечных тканей.
Косвенное перепрограммирование клеток (ILC)
Разработан метод при котором соматические клетки переходят в промежуточное пластическое состояние- частично перепрограммированные ИПСК (pre-iPSC), индуцированное кратковременным воздействием перепрограммирующих факторов, а затем дифференцируются с помощью специально разработанной химической среды (искусственной ниши). Предполагается, что этот новый метод может быть более эффективным и безопасным, так как он, по мнению его авторов, не вызывает опухоли или другие нежелательные генетические изменения, и при этом позволяет получать требуемые клетки быстрее и c гораздо большим выходом по сравнению с другими методами. Тем не менее, безопасность этих клеток все же сомнительна — учитывая то, что преобразование из пре-ИПСК опирается на использование условий перепрограммирования в ИПСК, и нельзя исключить что часть клеток может все же приобрести плюрипотентные свойства(если они не прекратят процесса де-дифференцировки in vitro или в связи с дальнейшей де-дифференцировкой in vivo).
Перепрограммирование воздействием на гликопротеин наружней мембраны
Общей особенностью, взятых из разных источников, плюрипотентных стволовых клеток, отличающей их от большинства (исключение составляют лейкоциты) неплюрипотентных клеток, является особый характер гликозилирования белков их наружней мембраны. Расположенные на поверхности стволовых клеток гликаны быстро реагируют на изменения в состоянии клетки и поэтому идеально подходят в качестве маркеров для выявления изменений в клеточных популяциях. Многие широко используемые маркеры стволовых клеток (в том числе SSEA-3, SSEA-4, TRA 1-60, и Тра 1-81.) являются гликанами клеточной поверхности. Так, например, гликопротеин подокаликсин (podocalyxin) расположен исключительно только на недифференцированных клетках человека (ИПСК и ЭСК), но не на поверхности дифференцированных соматических клеток, что позволяет отделить эти клетки с помощью лектина BC2L-C из Burkholderia cenocepacia (rBC2LCN). Suila Хели с соавт. полагают, что у стволовых клеток человека внеклеточные о-GlcNAc и о-LacNAc, выполняют решающую роль в тонкой настройке сигнального пути Notch — высококонсервативной системы клеточной сигнализации, от которой зависят судьбы стволовых клеток, их дифференцировка, лево- и правосторонняя асимметрия, апоптоз, и пролиферация (см. обзоры:)
Очевидно, изменения характера гликозилирования белков наружней мембраны являются маркерами состояния клетки каким-то образом связанными с плюрипотенцией и дифференцировкой. Причем «сдвиг» в характере гликозилирования, по всей видимости, это не просто результат инициализации экспрессии генов, а механизм играющий роль важного регулятора группы генов, вовлеченных в приобретении и поддержании недифференцированного состояния. Так, показано, что активация гликопротеина ACA, связывающего гликозилфосфатидилинозитол на поверхности прогениторных клеток периферической крови человека, посредством сигнального каскада PI3K/Akt/mTor/PTEN индуцирует повышение экспрессии генов Wnt, Notch1, Bmi-1 и HoxB4, а также способствует образованию и самообновлению популяции гемопоэтических стволовых клеток. Более того показано, что индуцируемая посредством ACA-зависимого сигнального пути дедифференцировка прогениторных клеток, приводит к образованию ACA-индуцированных плюрипотентных стволовых клеток, способных дифференцироваться in vitro в клетки всех трёх зародышевых листков.. Изучение избирательно связывающих гликопротеины лектинов, на предмет их способности поддерживать культуру плюрипотентных стволовых клеток человека, привело к открытию лектина эритрина кристагалли (Erythrina Cristagalli — ЕСА) способного служить в качестве простой и высокоэффективной матрицы для культивации человеческих плюрипотентных стволовых клеток
Индуцированные стрессом стволовые клетки (ИССК)
Клетки STAP (Stimulus-triggered acquisition of pluripotency)
В 2014 году группа японских исследователей опубликовала в статью в журнале Nature, где было заявлено открытие нового способа быстрого перепрограммирования соматических клеток млекопитающих в плюрипотентные клетки — так называемые клетки STAP в ответ на действие сильных внешних раздражителей, таких как временное повышение кислотности окружающей среды. Однако другим исследователям не удалось воспроизвести эти результаты. Впоследствии материал о клетках STAP был отозван журналом Nature как ошибочный, один из соавторов работы покончил жизнь самоубийством, а сами работы по этому направлению были прекращены.
Перепрограммирование индуцированное физическим воздействием
Плюрипотентные клетки содержат E-кадгерин, который при дифференцировке заменяется на N-кадгерин. Уникальной особенностью Е-кадгерина, помимо того, что он ответственен за межклеточную адгезию, является способность регулировать сигнальные пути клетки и заменять фактор Oct4 при индукции плюрипотентности. Фибробласты в которых подавлен синтез E-кадгерина не могут перепрограммироваться. Во время перепрограммирования, N-кадгерин может заменять функции E-кадгерина, что предположительно указывает на необходимость адгезии для перепрограммирования. Однако, согласно Гуаньнань Су с соавт., формирование в культуре клеток 3D сфер, в связи с вынужденным ростом клеток на поверхности с низкой связывающей способностью, иногда приводит к репрограммированию клеток. В качестве примера они показали, что нервные клетки-предшественники могут быть получены непосредственно из фибробластов путём физического воздействия, без введения экзогенных перепрограммирующих факторов. Ранее подобные сферы были получены в опытах с фибробластами мыши с мутацией инактивирующей ген-супрессор опухолей ретинобластомы — RB1, белка без которого клетки теряют способность к старым контактам и контактному ингибированию пролиферации в результате чего выходят за пределы колонии и образуют сферы где доминируют новые межклеточные контакты, по всей видимости, и вызывающие самопроизвольное перепрограммирование в тератомоподобные стволовые клетки.
Физические сигналы, в виде параллельных микродорожек на поверхности подложки для культивации клеток, могут заменить действие низкомолекулярных эпигенетических модификаторов и значительно повысить эффективность перепрограммирования. Механизм основан на механомодуляции изменяющей морфологию и эпигенетическое состояние клеток. В частности, по мнению авторов исследования: «снижение активности гистоновой дезацетилазы и повышение экспрессии WD повторяющего домена 5 (WDR5)-субъединицы H3 метилтрансферазы вызванное поверхностью с микродорожками приводит к увеличению ацетилирования и метилирования гистона Н3». Аналогичное действие на клетки оказывали нановолоконные подложки с выровненной ориентацией волокон.

Важным биофизическим фактором влияющим на дифференцировку клеток является жесткость подложки. Например, мягкие субстраты способствуют образованию из ЭСК, по BMP4-зависимому пути, нейроэпителиальных клеток, в то же время предотвращают дифференцировку в клетки нервного гребня. Исследования показали, что в этом механизме задействованы механочувствительное Smad фосфорилирование и ядерно-цитоплазматические перемещения, зависящие от регулируемой жесткостью подложки активности Hippo/YAP1 и сократительной способности комплекса актомиозин-цитоскелет.
Помогает клетке преобразовывать механические раздражители в электрические и биохимические сигналы белок регулирующий открытие ионного канала Са++ названный Пьезо1 (Piezo1), который активируется натяжением мембраны. В зависимости от липидного состава мембран придающего ей жесткость или мягкость меняется и способность Пьезо реагировать на механические стимулы
Механизмы механомодуляции см. в обзорах:
Разработан метод, который превращает соматические клетки в стволовые клетки «сжимая» их с помощью 3D микроокружения состоящего из специально подобранного геля, что открывает путь для крупномасштабного производства стволовых клеток для медицинских целей.
Как отмечено выше, в процессе перепрограммирования клетки морфологически изменяются, что приводит к изменению их способности к адгезии. Эти характерные различия в адгезии позволили разработать процесс выделения плюрипотентных стволовых клеток с помощью микрожидкостныхустройств. Преимуществом этого метода является то что: разделение занимает менее 10 минут, при этом удается получить более чем на 95 % чистую культуру ИПСК клеток, причем выживаемость клеток больше 80 % и полученные клетки сохраняют нормальные транскрипционный профиль, потенциал дифференцировки и кариотип.
Индуцированные нервные стволовые клетки (иНСК)
Центральная нервная система млекопитающих имеет крайне ограниченные возможности для регенерации. Поэтому для лечения многих нервных расстройств (таких как: инсульт, болезнь Паркинсона, болезнь Альцгеймера, боковой амиотрофический склероз и т. д.) требуются нервные стволовые клетки, автологичным источником которых могут стать иНСК пациента. В ряде новейших публикаций описано прямое преобразование соматических клеток в индуцированные нервные стволовые клетки.
Так, например, предшественники нервных клеток можно получить прямым преобразованием и без введения экзогенных транскрипционных факторов, пользуясь только химическим коктейлем. Эти клетки, называемые ciNPCs (chemical-induced neural progenitor cells) можно к примеру получить из фибробластов кончика хвоста мыши или мочевыводящих соматических клеток человека, используя для этого коктейль состоящий из:
- ингибитора HDAC (в качестве такового можно использовать либо вальпроевую кислоту), либо бутират натрия, либо трихостатин А;
- ингибитора GSK-3 (в качестве такового можно использовать либо CHIR99021, либо карбонат или хлорид лития);
- ингибитора сигнальных путей TGF бета (либо RepSox, либо SB-431542, либо Tranilast) и поместив клетки в условия гипоксии.
Аналогичным образом без введения экзогенных транскрипционных факторов, пользуясь только химическим коктейлем можно получить Шванновские клетки. По некоторым данным, в принципе, возможно, преобразовать трансплантированные в мозг мыши фибробласты и астроциты человека, спроектированные методами генной инженерии на выработку факторов (Ascl11, Brn2a и Myt1l) индуцирующих их перепрограммирование в нейроны, активируя после трансплантации соответствующие гены с помощью активатора добавленного к питьевой воде животных. Было также показано, что in situ эндогенные астроциты мыши могут быть напрямую преобразованы в функциональные нейроны, способные участвовать в формировании нейросетей. Важно отметить, что в отличие от ИПСК, полученные таким образом клетки не пролиферируют, а значит более безопасны. Наблюдения за подвергшимися этой процедуре мышами в течение года, не выявили у них признаков образования опухоли. Те же исследователи превратили астроциты спинного мозга в прогениторные клетки, называемые нейробластами, которые способны дифференцироваться в нейроны при поврежденнии спинного мозга. В то время как нейроны взрослого человека обычно не в состоянии регенерировать после травмы спинного мозга, нейроны, полученные из человеческих ИПСК, после трансплантации крысам с травмами спинного мозга продемонстрировали значительный рост по всей длине центральной нервной системы животных. Важно отметить, что в эксперименте были использованы ИПСК полученные из клеток кожи, взятых от 86-летнего мужчины. Авторы исследования продемонстрировали, что полученные из ИПСК омоложенные нейроны способны прожить в костном мозге крысы не менее трёх месяцев и в течение этого срока не образовывали опухолей. К сожалению, такая клеточная терапия не привела к излечению крысы от паралича.
Inoue и его коллеги трансплантировали глиальные нервные клетки-предшественники, полученные из человеческих иПСК в поясничный отдел спинного мозга мышей с моделью бокового амиотрофического склероза (БАС). Трансплантированные клетки дифференцировались в астроциты и продлевали жизнь мышей с БАС. Очевидно ИПСК могут стать перспективным ресурсом для трансплантационной терапии БАС.
Разработана технология для прямого преобразование фибробластов в функциональные астроциты с помощью транскрипционных факторов NFIA (Nuclear factor 1 A), NFIB (Nuclear factor 1 B) и SOX9
Как показано в обзоре Бельмонто с соавт. способы прямого преобразования соматических клеток в индуцированные нервные стволовые клетки отличаются по своим методическим подходам. Какой из этих подходов окажется наиболее приемлемым для клиники покажут исследования.
Прогениторные клетки олигодендроцитов (ПКОД)
Без миелиновой оболочки, выполняющей роль изоляции волокон нейронной сети, сигналы посланные по нервам быстро затухают. Поэтому при заболеваниях, связанных с потерей миелиновой оболочки, таких как рассеянный склероз, наблюдается снижение интеллекта, парез, атаксия туловища и конечностей, нарушения зрения, потеря чувствительности и ряд других неврологических симптомов. Перспективным подходом к лечению подобных заболеваний является трансплантация клеток-предшественников олигодендроцитов (ПКОД), способных заново создать миелиновую оболочку вокруг пораженных нервных клеток. Для такой терапии необходимо иметь доступный источник этих клеток. Основу для решения этой проблемы заложил метод прямого преобразования фибробластов мышей и крыс в олигодендроглиальные стволовые клетки индуцированного путём принудительной гиперэкспрессии восьми или всего трёх транскрипционных факторов Sox10, Olig2 и Zfp536. Показано, что аутологичная клеточная терапия с использованием полученных in vitro из ИПСК пациента клеток-предшественниц олигодендроцитов приводит к миелинизации in vivo, что свидетельствует о функциональности этих человеческих клеток в организме мыши и об открывшейся перспективе их использования в клинике.
Индуцированные кардиомиоциты (иКМ)
Одной из наиболее актуальных задач клинической науки нынешнего столетия является развитие терапевтических стратегий, способных обратить вспять прогрессирование сердечной недостаточности — основной причины инвалидности и смертности населения. Большие надежды в этом плане возлагаются на методы клеточной терапии, которые могли бы предотвратить образование соединительной рубцовой ткани вместо мышечной. Простейшим подходом к решению этой задачи могло бы быть перепрограммирование сердечных фибробластов непосредственно в организме путём доставки в сердце факторов транскрипции или микроРНК. Была предпринята попытка перепрограммировать сердечные фибробласты в кардиомиоцит-подобные клетки in vivo путём гиперэкспрессии в них факторов транскрипции Gata4, Mef2c и Tbx5 (GMT). В случае удачи, такой поход позволил бы превращать рубцовую ткань в мышечную непосредственно в сердце, без необходимости клеточной трансплантации. Эффективность такого перепрограммирования оказалась очень низкой, а фенотип полученных кардиомиоцитов существенно отличался от фенотипа нормальных зрелых кардиомиоцитов. Результатом чего явилась низкая выживаемость перепрограммированных клеток. Позднее в опытах in vitro фенотип удалось несколько исправить (добавлением ESRRG, MESP1, Myocardin, ZFPM2 и TGF-β), но эффективность перепрограммирования осталась низкой. Поднять эффективность перепрограммирования in vivo позволяют неинтегрирующиеся векторы вируса Сендай, с вектором факторов перепрограммирования Gata4, Mef2c, и Tbx5
Определённые успехи наметились в методах получения и выращивания большого количества кардиомиоцитов in vitro. Так, например, удалось с высокой степенью эффективности получить из ИПСК человека прогениторные сердечные клетки способные, при трансплантации их в сердечную мышцу, снизить её перерождение в рубцовую ткань после инфаркта. С помощью малых молекул и путём активации синтеза β-катенина или же ингибирования синтеза Wnt в ИПСК человека in vitro, удалось повысить эффективность получения кардиомиоцитов до 80 %.
Возможно, что в будущем удастся заменить искусственные электрокардиостимуляторы, необходимые людям с медленным или нерегулярным сердцебиением, на биологический кардиостимулятор (пейсмекер) из индуцированных стволовых клеток. Надежду на это вселяют эксперименты в которых поросятам делали инъекцию индуцированных сердечных клеток, способных синхронизировать ритм сердцебиения. Более того, при ишемической кардиомиопатии, вызванной смоделированным на мышах инфарктом миокарда, трансплантация ИПСК способствовала синхронизации поврежденных желудочков сердца, улучшая их проводимость и сократимость за счет активации процессов восстановления. Перепрограммированием соматических клеток in vivo с помощью эмбрионального фактора транскрипции T-box 18 (TBX18) можно преобразовать кардиомиоциты в клетки пейсмекера. Это открытие открывает возможность легко и быстро вылечить пациентов зависящих от кардиостимулятора. Перенос гена TBX18 In situ с помощью инъекции его аденовирусного носителя позволяет создать в месте инъекции естественный источник биологического водителя ритма уже через 2-3 дня после введения. При этом пока не наблюдалось возникновения опухолей или каких-либо нарушений в деятельности сердца. Таким образом, минимально инвазивный перенос генов TBX18 может рассматриваться как перспективный метод лечения больных с блокадой сердца, который в будущем, очевидно, заменит лечение искусственными кардиостимуляторами.
Создан коктейль для прямой (без прохождения через плюрипотентное состояние) трансдифференцировки, состоящий из четырёх низкомолекулярных компонентов (SB431542 (ингибитора ALK4/5/7), CHIR99021 (ингибитора GSK3), парната (ингибитора LSD1/KDM1, называемого также транилципромином), и форсколина (активатора аденилатциклазы)). Этот коктейль позволил с высокой эффективностью превратить фибробласты мыши в клетки сердечной мышцы с помощью всего одного фактора транскрипции — Oct4. Полученные таким способом индуцированные кардиомиоциты спонтанно сокращались. Методом прямой трансдифференциации без использования генетических векторов, то есть чисто фармакологически, с помощью коктейля из девяти компонентов, удалось получить с выходом 97 % из фибробластов кожи бьющиеся химически индуцированные кардиомиоцит-подобные клетки (ciCMs), которые почти не отличались от человеческих кардиомиоцитов по данным исследования их транскриптома, эпигенетически и по электрофизиологическим параметрам. Более того, при трансплантации в сердце мыши с инфарктом, обработанные этим коктейлем фибробласты превращались в выглядевшие здоровыми клетки сердечной мышцы. Предпринята успешная попытка противостоять постинфарктному фиброзу (перерождению сердечной мышцы в соединительную ткань с образованием рубца) с помощью химического перепрограммирования in vivo сердечных фибробластов в кардиомиоциты.
Лу с соавторами создали биоинженерную конструкцию сердца путём заселения очищенного от клеток (децеллюларизованного) сердца мыши мультипотентными сердечно-сосудистыми прогениторными клетками, полученными из ИПСК человека. Они обнаружили, что мультипотентные сердечно-сосудистые прогениторные клетки направленно мигрируют в соответствии с архитектурой сердца, а прибыв на место, размножаются и дифференцируются в кардиомиоциты, клетки гладких мышц и эндотелиальные клетки, как это необходимо для восстановления утраченной структуры сердца. Очевидно, что внеклеточный матрикс сердца мыши (оставшаяся после удаления клеток мыши подложка сердца) может посылать сигналы мультипотентным сердечно-сосудистым прогениторным клеткам человека, необходимые для их навигации и превращения в специализированные клетки, обеспечивающие нормальную работу сердца. Через 20 дней после перфузии сердца средой содержащей факторы роста, оно, после электростимуляции, начинало биться с темпом 40-50 ударов в минуту и реагировало на медикаменты.
Созревание кардиомиоцитов in vivo
Кардиомиоциты получаемые из ИПСК отличаются от взрослых соматических клеток и остаются незрелыми при культивировании в чашках Петри. Японским ученым удалось добиться созревания этих клеток. Для этого они на месяц поместили незрелылые клетки кардиомиоцитов в сердце новорожденной мыши для дозревания.
Омоложение мышечных стволовых клеток
Пожилые люди нередко страдают от прогрессирующей дистрофии и слабости мышц, что отчасти связано с повышенной активностью сигнальных путей p38α и p38β митоген-активированных протеин киназ в стареющих мышечных стволовых клетках. Подвергнув такие стволовые клетки непродолжительному воздействию SB202190 — ингибитора p38α и p38β — в сочетании с культивированием на мягкой подложке из гидрогеля, можно быстро омолодить их и размножить. Более того, после имплантации в организм такие омоложенные клетки способны повысить силу старых мышц. Восстановить способность сателлитных стволовых клеток к регенерации можно и подавив синтез на гене p16INK4a (называемом также Cdkn2a).
Миогенные предшественники, которые могут быть использованы для моделирования болезней или клеточной терапии скелетных мышц, могут быть также получены непосредственно из ИПСК с помощью свободно-плавающей шаровой культуры (EZ сфер) в культуральной среде, содержащей высокие концентрации (100 нг / мл) фактора роста фибробластов-2 (FGF-2) и эпидермального фактора роста.
|
| |
|---|---|
| Так выглядят фибробласты меченые зеленым флуоресценцентным красителем | |
Индуцированные гепатоциты
Получение клеток печени из ИПСК
Гепатоциты человека имеют очень ограниченную способность к восстановлению после повреждений печени. Поэтому трансплантация печени нередко является единственным способом лечения таких болезней как цирроз. Клеточная терапия печени затрудняется тем, что культура гепатоцитов плохо размножается in vitro. Поэтому удобнее размножить клетки в виде ИПСК, и только затем превратить их в гепатоциты. Разработано несколько способов получения гепатоцитов из ИПСК Так, например для очистки и размножения самообновляющихся гепатобласт-подобных клеток из человеческих плюрипотентных стволовых клеток (ЭСК/ИПСК), их культивировали на чашках покрытых человеческим ламинином-111 в течение более 3 месяцев, после чего они подобно овальным клеткам печени были способны дифференцироваться в гепатоцит-подобные клетки, а также в клетки желчных путей — холангиоцит-подобные клетки. Было показано, что такие гепатобласт-подобные клетки могут интегрироваться в паренхиму печени мыши. Предполагается, что, благодаря подавлению неблагоприятных генных регуляторных сетей при культивировании на поверхности покрытой ламинином, гепатоциты имеют большое сходстве с взрослыми гепатоцитами и могут быть использованы для скрининга лекарственных средств, а также в качестве источника клеток для регенеративной терапии печени.
В 2010 году была продемонстрирована возможность индуцировать полученные из жировой ткани стромальные клетки (ASC) в клетки похожие по ряду функций на гепатоциты человека, способные прижиться в поврежденной токсинами печени мыши. Позднее был разработан быстрый (до десяти дней) и эффективный (с выходом более 50 процентов) способ превращать клетки полученные путём липосакции в клетки печени. Клетки, полученные из собственных клеток человека с помощью этой новой методики, превращаются в клетки печени без промежуточной фазы плюрипотентных клеток и, очевидно, не образуют опухоли. В печени они формируют многоклеточные структуры необходимые для образование желчных протоков. Особенностью этой методики является культивирование адипоцитов в жидкой суспензии, в которой они образуют сфероиды
Обнаруженная у совместной культуры гепатоцитов, полученных из ИПСК, с эндотелиальными (для образования сосудов) и мезенхимальными (для образования поддерживающего внеклеточного матрикса) клетками, способность к самоорганизации (самосборке) в трёхмерные шарообразные структуры, представляющие собой зачаток печени позволяет надеяться, что в будущем трансплантологам: не надо будет искать и ждать донора, больному будут пересаживать зачаток нужного органа, полученный из его же собственных клеток, и этот зачаток будет уже на месте дорастать до нужных размеров. Эта методика позволяет использовать клетки всего одной мыши для предварительной проверки 1.000 лекарственных препаратов на их пригодность для лечения болезней печени, что открывает новые возможности для медицинских исследований и проверки безопасности лекарств.
Методы получения гепатоцитов без использования ИПСК
Для получения гепатоцитов из человеческих фибробластов не обязательно вначале получить ИПСК. Используя небольшие молекулы можно добиться прямого перехода фибробластов в индуцированные мультипотентные прогениторные клетки (iMPC) из которых затем образуются сначала прогениторные клетки эндодермы, а затем гепатоциты. После трансплантации мышкам с иммунодефицитом и смоделированным поражением печени, клетки iMPC интенсивно размножаются и приобретают функциональные способности характерные для взрослых гепатоцитов. Важно отметить, что при этом не наблюдалось образование опухолей, потому что клетки не проходили через стадию плюрипотентного состояния. С помощью инфицирования лентивирусами, вызывающими экспрессию генов FOXA3, HNF1A и HNF4A, удалось осуществить прямое преобразование фибробластов человека во взрослые гепатоцито-подобные клетки, которые могут быть размножены в культуре, а затем использованы для лечения острой печеночной недостаточности и метаболической болезни печени..
Инактивация сигнального пути Hippo in vivo с высокой эффективностью приводит к дедифференцировке взрослых гепатоцитов в клетки, несущие характеристики прогениторных клеток. Эти клетки-предшественники продемонстрировали способность к самообновлению и смогли прижиться в печени. Эти данные продемонстрировали беспрецедентный уровень фенотипической пластичности зрелых гепатоцитов
Коктейль из малых молекул, Y-27632, A-83-01 и CHIR99021, может превратить зрелые гепатоциты крысы и мыши in vitro в пролиферативные бипотентные клетки — CLiPs (chemically induced liver progenitors — химически индуцированные клетки-предшественники печени). CLIPS могут дифференцироваться как в зрелые гепатоциты, так и в эпителиальные клетки желчных протоков, которые могут образовывать функциональные структуры протоков. При длительном культивировании CLIPS не теряют свою пролиферативную активность и способность дифференциации в клетки печени, и могут заселять хронически пораженные ткани печени.
Подробнее см. обзор:

.
Клетки продуцирующие инсулин
Осложнения сахарного диабета, такие как сердечно-сосудистые заболевания, ретинопатия, невропатия, нефропатия и заболевания периферического кровообращения обусловлены дисрегуляцией сахара в крови из-за недостаточной продукции инсулина панкреатическими бета-клетками и при отсутствии адекватного лечения могут привести к летальному исходу. Одним из перспективных подходов к лечению диабета является трансплантация β-клеток, источником которых могли бы стать плюрипотентные стволовые клетки, (в том числе ЭСК и ИПСК). Однако β-клетки, получаемые из плюрипотентных стволовых клеток, имеют фенотип характерный для функционально незрелых β-клеток эмбрионального типа и отличаются от взрослых β-клеток повышенным уровнем базальной секреции глюкозы и отсутствием способности реагировать на сигналы стимуляции её синтеза (что подтверждают и результаты секвенирования РНК транскриптов).
Избыточная экспрессия комбинации трёх транскрипционных факторов (PDX1, NGN3 и MAFA) называемой PNM, способна привести к трансформации некоторых видов клеток в состояние подобное β-клеткам. Оказалось, что наиболее подходящим и доступным источником для перепрограммирования в инсулин-продуцирующие клетки, является эпителий кишечника. Под действием PMN трёхмерная культура зачатков органа (так называемые органоиды) стимулирует превращение эпителиальных клеток кишечника в β-подобные клетки, которые можно использовать для трансплантации.
Биоинженерия клеток кровеносных сосудов
Кровеносные сосуды образуют обширные сети, которые в течение всей жизни обеспечивают клетки организма питательными веществами и кислородом. Когда кровеносные сосуды становятся старше, их структура и функции, нередко, отклоняются от нормы, способствуя тем самым многочисленным возрастным заболеваниям, таким как: инфаркт миокарда, ишемический инсульт и атеросклероз артерий, питающих сердце, мозг и нижние конечности. Поэтому, важной задачей является стимулирование роста сосудов для обеспечения циркуляции, чтобы предотвратить обострение этих заболеваний. Одним из способов стимулирования роста сосудов является имплантация индуцированных прогениторных клеток эндотелия (иПЭк). Так, например, с помощью иПЭк, полученных путём частичного репрограммирования клеток эндотелия, удалось добиться увеличения коронарного кровотока и по данным эхокардиографии улучшить функционирование сердца. Стволовые клетки, извлеченные из жировой ткани после липосакции можно превратить в прогениторные гладкие мышечные клетки (иПГМк), участвующие в формировании артерий и вен. Это клетки могут быть использованы для создания кровеносных сосудов, необходимых для замены неисправных артерий сердца. Так, например, обнаружено, что с помощью культуры ИПСК человека в сочетании с селекцией с помощью трёх маркеров: CD34 (поверхностного гликофосфопротеина ранних эмбриональных фибробластов), NP1 (рецептора — нейрофилин1) и KDR (киназы содержащей домен рецептора), удалось получить эндотелиальные клетки, которые после трансплантации мышам образовали in vivo стабильные функциональные кровеносные сосуды, работавшие на протяжении по меньшей мере 280 дней..
При лечении инфаркта миокарда важно предотвратить образование фиброзных тканей шрама и стимулировать регенерацию. Достичь этого in vivo можно применив паракринные факторы способные изменить направление дифференцировки сердечных стволовых клеток предшественников от специализации в фиброзную рубцовую ткань в сторону образования сердечно-сосудистой ткани. Например, на мышиной модели инфаркта миокарда, было показано, что однократная интрамиокардиальная инъекция мРНК фактора роста эндотелия сосудов (VEGF-A modRNA), синтетически модифицированной так чтобы предотвратить её деградацию организмом, приводит к длительному улучшению функции сердца, обусловленному перенаправлением дифференцировки эпикардиальных клеток-предшественников в сердечно-сосудистый тип клеток.
Мервин Иодер с соавт., описали метод для преобразования ИПСК человека в клетки, подобные эндотелиальным колониеобразующим клеткам пуповинной крови (CB-ECFCs). Полученные ими CB-ECFC-подобные клетки имели стабильный эндотелиальный фенотип, высокий пролиферативный потенциал и способность, при трансплантации мышкам, образовывать человеческие кровеносные сосуды, а также участвовать в регенерации сетчатки и конечностей мыши после ишемии. Важно отметить, что индуцированные CB-ECFC-подобные клетки практически не образуют тератомы.
Прямое перепрограммирование клеток взрослого организма в прогениторные нефроны (ИПН)
Взрослые клетки проксимальных канальцев почки могут быть непосредственно перепрограммированы в прогениторные нефроны эмбриональной почки, с использованием пула из шести генов кодирующих «инструктирующие» факторы транскрипции (SIX1, SIX2, OSR1, Eyes absent homolog 1(EYA1), Homeobox A11 (HOXA11) и Snail homolog 2 (SNAI2)). Возможность получения таких клеток позволит в будущем приступить к разработке методов клеточной терапии почечных заболеваний. Первые успехи на этом пути уже есть. Так, недавно было показано, что эмбриональные органоиды почки, сформированные путём самоорганизации из клеточной суспензии, после трансплантации их во взрослую почку крысы могут в ней прижиться.
Биоинженерия стволовых клеток крови
Одной из самых востребованных целей регенеративной медицины является возможность получения в неограниченном количестве гемопоэтических стволовых клеток, пригодных для трансплантации, из более зрелых или дифференцированных клеток крови, для того чтобы покрыть дефицит трансплантатов костного мозга. Чтобы запустить в фибробластах процессы гемопоэза в условиях in vitro достаточно всего четырёх транскрипционных факторов: Gata2, Gfi1b, cFos, и Etv6 . Их воздействие приводит к образованию клеток подобных эндотелиальным — прогениторным клеткам с последующим возникновением из них кроветворных клеток. Аналогичным образом, используя 6 транскрипционных факторов: Run1t1, Hlf, Lmo2, Prdm5, Pbx1, и Zfp37, а также ещё два фактора Mycn и Meis1 для повышения эффективности перепрограммирования, удалось получить гемопоэтические стволовые клетки из зрелых дифференцированных клеток крови.
См. также обзоры:
Эритроциты
Переливание эритроцитов необходимо для многих пациентов с травмами или гематологическими заболеваниями. Однако, на сегодняшний день, поставка эритроцитов зависит от добровольных доноров число которых недостаточно. Кроме того, переливание крови от доноров сопряжено с определённым риском из-за возможности передачи ряда инфекций. Решить эту проблему могло бы производство необходимых количеств эритроцитов вне организма. В принципе уже доказано, что эритроциты, полученные вне организма из мобилизованных CD34-позитивных клеток (CD это на англ. сокращенно кластер дифференцировки), способны выжить при переливании аутологичному реципиенту. Эритроциты, получаемые in vitro, как правило, содержат исключительно зародышевый гемоглобин (HbF), который непригоден для нормального функционирования эритроцитов во взрослом организме. Тем не менее, in vivo, после трансфузии полученных из ИПСК эритроидных прогениторных клеток содержащих ядро, наблюдалось переключение на синтез взрослой изоформы гемоглобина. Однако в этом случае возникает другая проблема: несмотря на то, что эритроциты не имеют ядер, и, следовательно, не могут образовывать опухоли, их непосредственные предшественники эритроидные прогениторные клетки ядром обладают и следовательно потенциально опасны. Созревание эритробластов в функционально зрелые эритроциты требует сложного процесса реорганизации, который заканчивается удалением ядра с образованием безъядерных эритроцитов. Увы, методы перепрограммирования клеток в настоящее время часто приводят к нарушению этих процессов энуклеации и поэтому использование эритроцитов или их непосредственных предшественников эритробластов для переливания ещё недостаточно защищено от возможности образования опухолей. Тем не менее Bouhassira и его коллеги обнаружили, что кратковременное воздействие цитокинов, благоприятствующих дифференцировке стволовых клеток в эритроидные, на CD34 позитивные клетки, до их размножения с последующей пролиферацией полученных предшественников, позволяет получать на порядок больший выход эритроидных клеток, чем наблюдалось ранее. И что самое главное: эти красные кровяные клетки имели те же изоформы глобина что и использованные в качестве источника CD34 позитивные клетки. Значительно повысить выход эритроидных клеток из ИПСК или же эритроцитов из человеческих гемопоэтических стволовых клеток позволяет подавление гена SH2B3 или его инактивация генным редактированием с помощью CRISPR/Cas9
Интересно также отметить, что важную роль в развитии нормальных клеток крови играет сигнальный путь рецептора арил-углеводородов (AhR) (который как было установлено, содействует и образованию раковых клеток). Активация AhR в человеческих гемопоэтических клетках-предшественницах (HPS) приводит к беспрецедентной пролиферации HPS, мегакариоцитов и клеток эритроидных линий..
Подробный обзор методов получения эритроцитов см. в
Тромбоциты
Тромбоциты играют важную роль в предотвращении кровоизлияния у больных с тромбоцитопенией или с тромбоцитемией. Серьезной проблемой для пациентов после повторных переливаний тромбоцитов является развитие иммунных реакций. Поэтому для клиники большое значение имеет возможность получения тромбоцитов, не содержащих HLA-антигены, вне организма и на средах, не содержащих сыворотки. Некоторых успехов в этом направлении добились Figueiredo с соавторами. Используя РНК-интерференцию для подавления синтеза β2-микроглобулина в CD34-положительных клетках, они сумели получить тромбоциты, в которых на 85 % было снижено содержание антигенов HLA.
Разработан метод получения тромбоцитов, который заключается в создании из ИПСК человека устойчивых иммортализованных линий клеток-предшественников мегакариоцитов (imMKCLs) путём доксициклин-зависимой гиперэкспрессии Bmi1 и BCL-XL. Полученные imMKCLs можно размножать и культивировать в течение длительного периода (4—5 месяцев), причем даже после криоконсервации. Прекращение сверхэкспрессии c-MYC, Bmi1 и Bcl-X L (путём удаления доксициклина из среды) заставляло эти клетки производить тромбоциты CD42b+, которые по большинству параметров не отличались от тромбоцитов крови.
Альтернативный подход получения мегакариоцитов, с высоким выходом (3 единицы (2.4 × 1011 тромбоцитов на единицу) тромбоцитов для переливания с одного миллиона клеток ИПСК) и с чистотой более 90 %, позволяет культивирование на среде без продуктов животного происхождения (а потому с достаточно определёнными, предсказуемыми условиями, что важно для получения надёжно воспроизводимых результатов). Для перепрограммирования использовалась лентивирусная трансдукция, приводящая к одновременной экзогенной экспрессии трех факторов транскрипции: GATA1, FLI1 и TAL1.
Обзор по проблемам связанным с производством тромбоцитов см.
Иммунные клетки
Вырабатываемый иммунной системой специализированный тип белых кровяных клеток, известный как цитотоксические Т-лимфоциты (CTL), способен распознать специфические маркеры на поверхности различных инфекционных или опухолевых клеток и уничтожить эти вредоносные клетки. Поэтому иммунотерапия с использованием антиген-специфических Т-клеток в будущем может быть использована для борьбы со многими видами рака и вирусных инфекций. Организм производит очень мало таких клеток и выделить их в количестве необходимом для терапии очень сложно. Потенциально эффективным подходом получения этих клеток для терапии может быть технология заключающаяся в том чтобы превратить зрелые CTL в ИПСК, которые обладают способностью к неограниченной пролиферации in vitro, размножить эти ИПСК до необходимого количества и затем провести их дифференцировку обратно в зрелые CTL. Ещё большие возможности обещает метод, который сочетает две технологии — 1. получение ИПСК и превращение их в Т-клетки, и 2. последующую их генетическую модификацию, с помощью технологии конструирования химерных рецепторов антигенов (CAR), позволяющую им распознавать раковые клетки-мишени по антигенам, в частности по CD19 — антигену, синтезируемому злокачественными В-клетками. По аналогичной технологии можно было бы создать распознающие белок PBP2A Т-клетки направленные против устойчивых к антибиотикам бактерий таких как метициллин-резистентный золотистый стафилококк.
Большой клинический потенциал в качестве адъюванта для иммунотерапии рака имеют инвариантные естественные киллеры T (INKT) — клетки, которые могут служить в качестве моста между врожденной и приобретенной иммунной системами. Они повышают противоопухолевую активность организма, производя гамма-интерферон (ИФН-γ). Предложен концептуальный метод использования INKT клеток, полученных из ИПСК, для терапии рака, который состоит из четырёх ступеней: (1) выделение минимального количества INKT клеток, (2) перепрограммирования этих INKT клеток в ИПСК, (3) размножение этих ИПСК в культуре и дифференцировка их обратно в INKT клетки и (4) инъекция, полученных INKT клеток, подопытным животным для терапии рака.
Для терапии могут быть использованы также дендритные клетки, которые участвуют в контроле Т-клеточного ответа. После инъекции они могут выжить достаточно долго, чтобы стимулировать антиген-специфические CTL, после чего могут быть полностью устранены. Неисчерпаемым источником для терапии вакцинами могут служить антиген-представляющие дендритные клетки, полученные из человеческих ИПСК или прямым перепрограммированием из фибробластов.
В-клетки способны к быстрой (за 2-3 дня) трансдифференцировке в макрофаги под воздействием транскрипционного фактора C / EBPα. Кроме того эффективность перепрограммирования В-клеток в ИПСК с помощью транскрипционных факторов Oct4, Sox2, Klf4 и Myc под воздействием C / EBPα возрастает 100-кратно и охватывает порядка 95 % популяции клеток. Интересно отметить, что с помощью C / EBPα можно преобразовать некоторые линии в-клеток лимфомы человека и лейкоза в макрофаг-подобные клетки уже не способные к дальнейшему онкогенезу.
Омоложение эпителиальных клеток тимуса
Тимус является органом, размеры которого существенно сокращаются с возрастом. Это сокращение является одной из основных причин того, что иммунная система с возрастом становится менее эффективной. Одним из центральных звеньев механизма возрастной инволюции тимуса является снижение синтеза транскрипционного фактора FOXN1. Клэр Блэкберн и её коллеги показали, что даже далеко зашедшая возрастная инволюция тимуса может быть обращена вспять путём принудительного усиления активности в эпителиальных клетках тимуса всего одного фактора транскрипции — FOXN1 с целью содействия омоложению, пролиферации и дифференцировки этих клеток в полностью функциональный эпителий. Более того они показали, что принудительная экспрессия Foxn1 позволяет перепрограммировать клетки кожи — фибробласты в функциональные эпителиальные клетки тимуса. Эти FOXN1-индуцированные эпителиальные клетки тимуса (iTECs) поддерживали эффективное развитие in vitro линий клеток CD4+ и CD8+ тимуса. Но, что самое главное, после трансплантации в почку мыши, iTECs собирались и образовывали полностью организованный и функциональный тимус, который содержал все подтипы эпителиальных клеток тимуса, необходимых для поддержки дифференцировки Т-клеток, в результате чего иммунная система реципиента пополнялась новыми Т-клетками. Это открытие можно считать первым примером выращивания органов из трансплантированных индуцированных стволовых клеток. В будущем этот метод может быть широко использован для повышения иммунной функции у пациентов путём трансплантации тимуса.
Индуцированные стволовые/стромальные клетки мезенхимы (ИМСК)
Благодаря своим способностям вызывать иммуносупрессию и способности к дифференцировке во многие типы мезенхимальных тканей, стволовые/стромальные клетки мезенхимы (МСК) интенсивно исследуются на предмет их применения для лечения сердца, почек, нервной ткани, суставов и регенерации костей, а также терапии воспалительных заболеваний и подавления реакции отторжения при трансплантации. МСК, как правило, получают путём болезненных, инвазивных процедур из взрослого костного мозга или жира, при этом выход очищенных МСК составляет всего лишь 0,001 % — 0,01 % от клеток костного мозга и 0,05 % от аспирата липосакции. На практике удобнее всего получать МСК из аспирата липосакции, при этом удаляют взрослые адипоциты, которые успели потерять способность к пролиферации. Между тем взрослые адипоциты легко можно выделить и подвергнуть дедифференцировке в так называемые дедифференцированные жировые клетки (ДДЖК), которые возвращают способность к пролиферации и мультипотентность. При соответствующих условиях культивирования in vitro или окружения in vivo ДДЖК могут дать начало адипогенным, остеогенным, хондрогенным или миогенным прогениторным клеткам, а также стимулировать неоваскуляризацию то есть проявляют те же свойства, что и МСК костного мозга. У пожилых пациентов, которые больше всего нуждаются в восстановлении тканей путём аутологичной клеточной терапии, с возрастом наблюдается резкое возрастное снижение количества и качества МСК и адипоцитов. Вместе с тем, известно, что ИПСК могут быть получены путём омоложения клеток даже от столетних людей. Поэтому ИПСК, которые можно получить перепрограммированием клеток из тканей пациента и затем практически неограниченно размножать in vitro, может стать удобным источником омоложенных МСК. Как показали опыты на мышах с моделью воспалительных заболеваний кишечника таких как Болезнь Крона и Язвенный колит, молоденькие ИМСК могут быть успешно использованы для лечения даже лекарственно-рефрактерных форм подобных воспалительных заболеваний.
Chen с соавт. обнаружили, что воздействуя на ИПСК человека препаратом SB-431542 можно достаточно быстро получить однородную культуру клеток ИМСК, которые по свойствам мало чем отличаются от молодых МСК. По мнению авторов статьи такие ИМСК не обладают способностью к образованию тератом и имеют стабильный кариотип, а поэтому могут быть использованы для терапии В настоящее время пока мало данных об эффективности и долгосрочной безопасности полученных этим методом ИМСК in vivo. Известно только что ИМСК могут быть использованы в клинике для лечении периодонтита и разработки методов ортопедии
Важную роль в инициировании и ускорении молекулярной программы, которая приводит к дифференциации ИМСК из ИПСК выполняет белок 2MSX2 (muscle segment homeobox 2). Генетическая делеция MSX2 ухудшает дифференцировку ИМСК из ИПСК. При использовании коктейля растворимых молекул эктопическая экспрессия MSX2 способствует образованию почти однородной популяции полностью функциональных ИМСК.
Разработан химический метод получения ИМСК из первичных фибробластов кожи человека с использованием шести химических ингибиторов (SP600125, SB202190, Go6983, Y-27632, PD0325901 и CHIR99021) с добавлением трёх факторов роста: трансформирующего фактора роста-β (TGF-β), основного фактора роста фибробластов (bFGF) и фактора подавления лейкемии (LIF). Этот химический коктейль преобразует человеческие фибробласты в ИМСК всего за 6 дней с эффективностью порядка 30-40 процентов.
Культуры мезенхимальных стволовых клеток человека могут быть использованы in vitro для массового производства экзосом, которые, как выяснилось, идеально подходят в качестве средства для доставки лекарств и для доставки в клетку-мишень факторов транскрипции или микроРНК индуцирующих перепрограммирование (дедифференцировку, дифференцировку или трансдифференцировку).
Индуцированные хондрогенные клетки (ИХОНК)
Хрящ соединительной ткани обеспечивает движение суставов без трения. Его дегенеративное перерождение в конечном счете, приводит к полной потере функции сустава на поздних стадиях остеоартрита. Единственным типом клеток в хряще являются хондроциты окруженные выделяемым ими внеклеточным матриксом. В настоящее время исследователи используют два способа восстановления хрящевой ткани:
- получением хондроцитов из плюрипотентных клеток (ЭСК/ИПСК).
- получением хондроцитов прямым преобразованием фибробластов человека непосредственно в индуцированные хондрогенные клетки, минуя промежуточную стадию плюрипотентных клеток, с помощью трёх факторов репрограммирования (с-Мус, KLF4, и SOX9).
Преимуществом первого метода является быстрое размножение культуры исходных клеток. Преимуществом второго — отсутствие в культуре плюрипотентных клеток, которые могли бы вызвать тератому. Клетки, полученные прямым перепрограммированием, синтезировали коллаген типа II. После имплантации в область поражения они смогли выжить и в течение не менее четырёх недель участвовали в образовании хрящевой ткани у мышей.
Ещё одним перспективным источником клеток для перепрограммирования являются мезенхимальные стволовые клетки, полученные из фолликулов человеческого волоса. и кератиноциты
Важно отметить, что происхождение соматических клеток используемых для перепрограммирования может оказывать влияние на эффективность перепрограммирования, функциональные свойства получаемых индуцированных стволовых клеток и способность к образованию опухолей.
При выборе источника для перепрограммирования, во внимание следует принимать тот факт, что ИПСК сохраняют эпигенетическую память о тканях из которых они произошли, и что это влияет на их способность к направленной дифференцировке Остаточная эпигенетическая память не обязательно проявляется на стадии плюрипотентности — ИПСК, полученные из разных тканей имеют надлежащую морфологию, в них активны гены характерные для плюрипотентности, и они способны дифференцироваться в ткани трёх эмбриональных слоев как in vitro, так и in vivo. Однако, эта эпигенетическая память может проявиться позже, во время повторной дифференцировки в специфические типы клеток, которая требует активации локусов, сохранивших элементы остаточной эпигенетической памяти.
Культуральная среда для плюрипотентных стволовых клеток свободная от питающих клеток и сыворотки
Для выращивания плюрипотентных стволовых клеток человека обычно используются так называемые питающие клетки (feeder cells) и сыворотка из эмбрионов быка (FBS). И то и другое является продуктами животного происхождения и может изменять свойства от партии к партии, что затрудняет стандартизацию условий. Кроме того выращивание стволовых клеток на клетках другого человека или животных создает риск загрязнения патогенными микроорганизмами, которые могут стать источником болезни для пациента после клеточной терапии.. Поэтому компоненты животного происхождения требуют дорогостоящего контроля на качество, и их свободу от патогенов, полиаминоксидазы и антигенов. Для замены питающих клеток используются различные подложки, такие как: Matrigel, CELLstart, рекомбинантные белки и синтетические полимеры такие как Synthemax (см. обзор к статье.
Известно, что важную роль в адгезии клеток друг к другу и к внеклеточному матриксу играет тримерный белок ламинин. Было найдено что ламинин-511, названный так за то, что он содержит α5 , β1 и γ1 цепи, если его нанести на дно чашки Петри способен поддерживать стабильную культуру ЭСК или ИПСК. На основе этого открытия была разработана стандартная методика для длительного культивирования ЭСК и ИПСК человека в чашках покрытых rLN511E8 — рекомбинантным фрагментом ламинина −511 и с безсывороточной средой StemFit™. Аналогичная методика но с использованием ламинина-521 и E-кадгерина позволила клонировать in vitro эмбриональные стволовые клетки без необходимости использовать ингибиторы ROCK (англ. Rho-associated protein kinase). Интересно было бы применить её и для ИПСК.
В стадии разработки находится также очень дешёвая подложка из углеродных нанотрубок. Она позволит выращивать и проводить дифференцировку стволовых клеток в промышленных масштабах. По заверениям авторов изобретения, изменяя условия изготовления этой подложки, можно так изменить её свойства, что она будет влиять на способность выращиваемых клеток к адгезии, на их пролиферацию и морфологию образуемых клеточных колоний.
Для трёхмерного культивирования широко используют гидрогели, такие как, например, гидрогель для получения кардиомиоцитов в одну стадию
Способы доставки репрограммирующих факторов в ядро
Способы доставки можно подразделить на вирусные и невирусные, а также на те, что связаны с интеграцией векторов несущих репрограммирующие факторы в геном и действующие без интеграции.
(Значками отмечены свойства соответствующего вектора: (+)- Геномная интеграция происходит; (±)- интеграция происходит, но очень редко; (-)- вектор не интегрируется; (тр)- после интеграции векторная конструкция должна быть удалена транспозазой.)
Доставка вирусами
Чаще всего для доставки используются вирусные векторные системы. Вирусы используют свой врожденный механизм заражения клетки, что позволяет использовать их для доставки и внедрения кассеты генов необходимых для экспрессии репрограммирующих факторов В качестве вирусов для доставки генов обычно используют:
- Ретровирусы(+). Они в качестве генома содержат одноцепочечную молекулу РНК. С помощью обратной транскрипции на РНК вируса синтезируется линейная двухцепочечная ДНК которая затем интегрируется в двухцепочечную ДНК генома клетки-хозяина. Описан метод эффективного перепрограммирования клеток человека в ИПСК с помощью одного вектора содержащего четыре ТФ, в сочетании с коктейлем, содержащим три небольшие молекулы. Приведены прописи аналогичных методов.
- Лентивирусы(+). Они являются подклассом ретровирусов. В отличие от ретровирусных векторов, лентивирусные векторы могут заражать не только делящиеся клетки, но и находящиеся в покое терминально дифференцированные клетки. Удаляемая полицистронная кассета STEMCCA представляющая собой вырезаемый Cre-рекомбиназой перепрограммирующий лентивирусный вектор позволяет осуществлять свободное от трансгенов перепрограммирование взрослых фибробластов кожи человека в ИПСК.
- Вирус Сендай (-) — вирус из семейства Paramyxoviridae, содержащий одноцепочечную РНК. Вирус Сендай считается безопасным, потому что его генетический материал не включается в ДНК клетки, и от него достаточно легко избавиться инкубацией культуры клеток при повышенной температуре. Вирус от тепла погибает, тогда как трансформируемым клеткам такая обработка не вредит. Для получения ИПСК этим методом можно воспользоваться готовыми наборами. В отличие от ретровирусных и эписомных векторов, при перепрограммировании с использованием вируса Сендай пока не наблюдалось дефектных клонов, не способных к дифференцировке. Описание метода см.
- Венесуэльский вирус лошадиного энцефалита (VEE) (-) у которого структурные белки удалены, но все ещё присутствуют неструктурные белки, позволяет с помощью самореплицирующегося полицистронного репликона РНК внести в клетку однократной трансфекцией четыре перепрограммирующих фактора (OCT4, KLF4, SOX2 и либо c-MYC либо GLIS1) .
- Не интегрирующиеся аденовирусы (±). По мнению некоторых авторов, векторную кассету, после того как перепрограммирование достигнуто, можно удалить с помощью трансфекции мРНК Cre рекомбиназы, что якобы позволяет сочетать высокую эффективность вирусной доставки с достоинствами перепрограммированных клеток, свободных от остатков трансгенов, которые могут вызывать злокачественную трансформацию.
Доставка c помощью аденоассоциированного вирусного вектора в экзосоме
Аденоассоциированный вирусный вектор (AAV) может быть связан с экзосомами (exo-AAV), если вектор выделяют из культуральной среды клеток-продуцентов. Такой вектор более устойчив к нейтрализующим антителам по сравнению со стандартным AAV. Он более эффективен для трансфекции in vivo
Невирусная доставка векторов
По сравнению с вирусными векторами, не-вирусные векторы являются потенциально менее иммуногенными и их сравнительно легче использовать в условиях клиники.
Невирусным подходом является прямая доставка в клетку синтетической мРНК(-) кодирующей четыре канонических фактора Яманаки: KLF4, c-MYC, OCT4, и SOX2. Метод позволяет достичь высокой эффективности перепрограммирования, но технически сложен и сильно зависит от качества реактивов. Недавно он был модифицирован, что позволило сократить продолжительность процесса и число необходимых реагентов. Ещё более экономным в отношении стоимости перепрограммирования является метод получения ИПСК и последующей их дифференцировки с помощью микрожидкостного устройства в объёмах не превышающих микролитра и эффективностью в 50 раз большей чем при традиционном перепрограммировании путём доставки синтетических мРНК, кодирующих факторы транскрипции.
Для перепрограммирования in vivo очевидно подойдет доставка мРНК с помощью олиго (карбонат-b-α-амино эфиров) под общим названием CARTs (charge-altering releasable transporters), катионов, которые образуя комплекс с мРНК, защищают её и доставив в клетку высвобождают
Перепрограммирование мышечных клеток головастиков Xenopus в недифференцированные клетки может быть достигнуто In vivo с помощью ДНК(±) мыши кодирующей Oct4, Sox2, и Klf4 в условиях способствующих регенерации.
Привлекательным методом невирусной доставки генов, который позволяет эффективно встраивать желаемую ДНК в геном различных клеток является использование транспозонов — сложных структур содержащих дискретные куски ДНК, обладающие способностью изменять своё местоположение в геноме посредством механизма транспозиции (инсерции), вынуждающего его включиться или наоборот покинуть определённый участок генома. Транспозон состоит из инсерционных сегментов ДНК, которые могут перемещаться как единое целое, захватывая лежащие между ними гены. Описано несколько транспозонных систем пригодных для транспортировки генов в клетки млекопитающих. Это «Спящая красавица» — Sleeping Beauty (SB), SB100X, и Tol2 и PiggyBac (PB). Разработаны эффективные методики получения ИПСК мыши и человека введением в соматические клетки векторов на основе PiggyBac (тр) В том числе с кассетой из ДНК кодирующей 6 факторов (Oct4, c-Myc, Klf4, Sox2 плюс Rarg (retinoic acid receptor gamma — RAR-γ) и Lrh-1 (liver receptor homologue 1 известный также как Nr5a2)., что позволило сократить продолжительность перепрограммирования до 4-12 дней и поднять его эффективность до уровня сопоставимого с методами переноса ядер и слияния клеток.
Векторные системы на основе эписомных плазмид(±). Перепрограммирование основанное на использовании эписомных плазмид считается наиболее эффективным и безопасным так как не требует интеграции трансгенов в геном. Однако первоначально эффективность перепрограммирования этим методом была крайне низка (менее 0,0002 %.) . Значительно увеличить эффективность индукции ИПСК позволило использование комбинации плазмид, кодирующих OCT3/4, SOX2, KLF4, L-MYC, LIN28 и малой интерферирующей РНК для ингибирования гена TP53, кодирующего белок p53 (супрессор опухолей, ингибирующий перепрограммирование) в сочетании с Эпштейн-Барр ядерным антигеном 1 (EBNA1), который является белком вируса Эпштейна-Барр (ВЭБ), необходимым для амплификации эписомных векторов Разработаны полицистронные свободные от вирусов плазмиды (±), содержащие последовательности самовыщепляющегося 2A пептида (пептида, который взаимодействуя с рибосомой инициирует отделение от растущего полипротеина, вызвав «пропуск» последней пептидной связи на С-конце 2А) и постоянно активный промотор вируса энцефаломиокардита (CMV).
Рекомбинантные белки (-) представляют собой проникающие в ядро белки, полученные путём рекомбинации (в данном случае, присоединения последовательности, кодирующей поли-аргининовый домен трансдукции, к генам четырёх перепрограммирующих факторов: Oct4, Sox2, Klf4 и C-Myc, на участке соответствующем их С-концевой последовательности, и, кроме того, постоянно активного промотора вируса энцефаломиокардита) с последующим синтезом этих белков в тельцах включения бактерии E.coli. Выделенные из E. coli и очищенные рекомбинантные белки используются для репрограммирования без интеграции. Эффективность репрограммирования с помощью рекомбинантных белков ничтожно мала, но может быть существенно увеличена при воспалении вызываемом поли I:C (синтетическим аналогом двухцепочечной РНК).
Химически индуцированные плюрипотентные клетки (ХИПСК)
Используя для перепрограммирования исключительно малые молекулы, китайские ученые Дэн Хонгкуй с коллегами показали, что для перепрограммирования клеток достаточно эндогенных «мастер генов». Они индуцировали плюрипотентное состояние у взрослых клеток мыши, используя семь низкомолекулярных соединений. Эффективность метода оказалась достаточно высокой: она была в состоянии преобразовать 0,2 % взрослых клеток ткани в ИПСК, что сопоставимо с результатами получаемыми при использовании экзогенных «мастер генов» доставляемых вирусными векторами. Авторы отмечают, что полученные от мышей CiPSCs были на «100 % жизнеспособными и здоровы на протяжении, по меньшей мере, 6 месяцев наблюдения». В число этих семи низкомолекулярных соединений входили:
- вальпроевая кислота — жирная кислота с разветвленной цепью — ингибитор гистондеацетилазы, повышает эффективность перепрограммирования соматических мышиных эмбриональных фибробластов (примерно в 100 раз), способствуя активации генов плюрипотентности и репрессии генов линейной дифференцировки.
- ингибитор GSK3 — гликоген синтез киназы 3 (CHIR) (подробнее см.). Ингибирование GSK3 имитирует активацию сигнального пути Wnt в результате чего β-катенин избежав деградации входит в ядро, где активирует синтез ферментативной субъединицы теломеразы (TERT);
- ингибитор трансформирующего ростового фактора бета (E-616452). Низкомолекулярный ингибитор TGF-[бета] сигнализации в ходе перепрограммирования заменяет Sox2 активируя Nanog;
- ингибитор моноаминоксидазы антидепрессант транилципромин одновременно является ещё и ингибитором деметилазы которая избирательно удаляет метильные группы с лизина в положении Лиз4 или Лиз9 гистона H3. Действуя на хроматин транилципромин вызывает глобальное увеличение H3K4 метилирования и как следствие этого дерепрессию генов . Интересно отметить, что ингибиторы деметилаз обладают ещё и способностью подавлять избыточную пролиферацию клеток и таким образом подавлять развитие раковых опухолей
- Форсколин (forskolin) — препарат, активирующий аденилатциклазу — фермент, катализирующий превращение АТФ в цАМФ;
- ингибитор гидролазы S-аденозилгомоцистеина (под названием 3-деазанепланоцин (DZNep)), вещество вызывающее разрушение поликомб репрессорного комплекса PRC2 и ингибирующее метилирование гистонов.
- PD-0325901 — высокоизбирательный и сильнодействующий синтетический ингибитор митоген-активируемой протеинкиназы MEK, избирательно фосфорилирующей серин / треонин и остатки тирозина в активационной петле её субстратов. PD-0325901 проходит испытания в качестве противоопухолевого препарата.
Методы и составы «коктейлей» из малых молекул для химического воздействия на мышечную ткань с целью активации процессов образования функциональных сердечных, скелетных и гладкомышечных клеток и регенерации поврежденных тканей in situ можно найти в обзоре Джанга и Виллиамса. С механизмами действия малых молекул при перепрограммировании можно ознакомиться в обзорах.
Прямое перепрограммирование фибробластов через универсальный XEN этап
В 2015-м году система химического перепрограммирования была усовершенствована — разбита на три этапа с использованием разных «коктейлей» на разных этапах. Были найдены новые малые молекулы. Это позволило поднять выход перепрограммированных клеток почти в 1000 раз. Выяснилось, что поскольку начальный этап химического перепрограммирования переводит клетки фибробластов в стабильное XEN (extra-embryonic endoderm)-подобное состояние одинаковое для прямого перепрограммирование фибробластов в различные клетки, эти XEN-подобные клетки (которые можно размножать) могут служить универсальной платформой для создания различных желаемых типов клеток.
Краситель для выявления плюрипотентных клеток человека
Для безопасной терапии индуцированными стволовыми клетками необходимы несложные методы обнаружения и разрушения недифференцированных стволовых клеток. С этой целью широко используются антитела SSEA-4 и SSEA-5, а также антитела на антигены: антитело TRA-1-60 на трансмембранный гликопротеин подокаликсин (podocalyxin), Oct3/Oct4 и Nanog. Эти вещества как и большинство белков стоят недешево и быстро портятся. Гораздо более стабильным и дешёвым реагентом способным отличить плюрипотентные стволовые клетки от дифференцированных клеток оказался флуоресцентный краситель KP-1 (Kyoto probe 1), который избирательно красит в митохондриях альдегиддегидрогеназу 2 (ALDH2). Эта селективность KP-1 зависит от способности митохондрии удалить его с помощью транспортных белков множественной лекарственной устойчивости ABCB1 и ABCG2, экспрессия которых подавляется в плюрипотентных клетках человека и индуцируется после дифференцировки.
Фотодинамическая технология удаления ИПСК
Способность плюрипотентных клеток, и в частности ИПСК, избирательно окрашиваться красным красителем CD1 может быть использована для их селективного удаления из инкубационной среды, с помощью фотодинамической обработки светом, оставшихся недифференцированными ИПСК после проведения их дифференцировки в соматические клетки. Эта простая технология предварительной очистки позволяет резко снизить риск образования опухолей типа тератомы при трансплантации клеток полученных из ИПСК. Разработана также разновидность этой технологии. Для этого флуоресцентную пробу KP-1 (Kyoto probe 1) (которая обычными клетками выводится наружу белками ABCB1 и ABCG2, синтез которых в ИПСК подавлен) соединили с анти раковым препаратом SN38. Полученный препарат, названный «конъюгат 17» за 72 часа полностью удаляет оставшиеся недифференцированными ИПСК
Химическая технология удаления ИПСК
Чтобы избавиться от плюрипотентных клеток, которые могут вызвать образование терратомы, можно использовать синтетические фосфо-D-пептиды, поскольку известно что на поверхности ИПСК сверхэкспрессируются щелочные фосфатазы, которые вызывают дефосфорилирование фосфо-D-пептидов в гидрофобные пептиды, которые агрегируя вызывают гибель клеток.
Перспективы изучения индуцированных стволовых клеток для медицины
Ярким свидетельством значения индуцированных стволовых клеток для медицины будущего и всего человечества, стало присуждение Джону Гордону и Синъе Яманаке Нобелевской премии 2012 года по медицине. Бо́льшую часть от Нобелевской премии, а также от полученной им в 2013 году Премии за прорыв в области медицины в 3 млн долларов Синъя Яманака решил потратить на развитие своих исследований. В настоящее время индуцированные стволовые клетки используются главным образом для моделирования болезней, скрининга (селективного отбора) лекарств, проверки токсичности различных препаратов. Однако в ближайшие годы начнется их широкомасштабное использование для клеточной терапии и выращивания органов и их «запчастей» для трансплантации.
См. также
- Регенеративная медицина
- Клеточная трансплантология
- Выращивание органов
- Репрограммирование клеток
- Examples of in vitro transdifferentiation by lineage-instructive approach (англ.)
- Examples of in vitro transdifferentiation by initial epigenetic activation phase approach (англ.)
- Examples of in vivo transdifferentiation by lineage-instructive approach (англ.)
- Induced stem cells